|
|
|
В.В. Матвеев
Рассказ о необычных представлениях американского ученого Гильберта Линга
G. N. Ling. A Revolution in the Physiology of the Living Cell, Krieger Publishing Company, Malabar, Florida, 1992.
(Другая версия этой статьи опубликована в журнале "Химия и жизнь" № 8, 1994, с.42-47)
Содержание.
Против методологического принуждения
Четыре фундаментальных свойства клетки
Основной вопрос физиологии клетки
Предсказательная сила теории, или А-диски против мембран
Элементарная структурная ячейка клетки
Фундаментальные свойства клетки с революционных позиций
Энергетическое инакомыслие
Имеют ли теории право на исключения?
Контрреволюционные возражения
Истоки революции
Живительная сила науки
Литература
Порядок в наши дни есть обычно там, где ничего нет.
Он указывает на бедность.
Бертольт Брехт
Когда берешь в руки эту книгу, невольно вспоминаются строки из Маяковского о чувствах некоего буржуазного чиновника, взявшего в руки "краснокожую паспортину": "Берет - как бомбу, берет - как ежа, как бритву обоюдоострую..." и т.д. Да, приблизительно с такими чувствами должен брать в руки книгу Линга всякий, кто глубоко убежден, что в физиологии клетки царит полный порядок. В самом деле, какие еще чувства может испытывать специалист, когда узнает, что мембранные насосы - это всего лишь сомнительная гипотеза, а не реальность? Что плазматическая мембрана полупроницаема не благодаря системе отверстий в непроницаемой стене билипидного слоя, а совсем по другим причинам? Что в основе распределения между клеткой и средой кислорода, K+, аминокислот, углеводов и многого другого лежат не разные, а один и тот же механизм - сорбция на внутриклеточных структурах?
Все это выглядит как попрание устоев, как кощунство над современным знанием. (Впрочем, есть мнение, высказанное Б. Шоу, что многие великие истины были вначале кощунством). Поэтому моя задача рассказчика особенно трудна. Излагая "вздорные" идеи, невозможно просто козырять фактами, пусть голыми и упрямыми. Факты услужливы в любом виде и качестве. По этой причине, чтобы не пасть жертвой "фактического" лицемерия, необходимо начать с нескольких слов...
ПРОТИВ МЕТОДОЛОГИЧЕСКОГО ПРИНУЖДЕНИЯ.
Научный метод - это совокупность правил, управляющих деятельностью науки. Если работа исследователя осуществляется в соответствии с этими правилами, то она научна, если нет - ненаучна. Эта видимая простота осложняется тем проклятым обстоятельством, что критерии научности не всегда формулируются явно. Конкретная практика науки настолько богата и многообразна, что возникает потребность интерпретировать требования научного метода применительно к каждому конкретному случаю. А там, где интерпретация - без интуиции не обойтись. Из этой размытости понятия "научного метода" и рождается неистребимая гидра методологического принуждения, наиболее страшная метаморфоза которой - господствующее в науке общественное мнение.
Тираническая природа общепринятых представлений состоит в том, что на деле теории всегда давлеют над фактами, и в истории науки еще ни одному самому очевидному факту не удалось легко и просто развалить теорию, которой он противоречил самым вопиющим образом.
Верить легче, чем анализировать. Вера в определенные ценности принуждает нас самих по-своему трактовать совокупность правил, громко именуемых научной методологией. В итоге, ученый не только является жертвой методологического принуждения, но и сам всегда готов выступить в роли безжалостного палача, ревностного хранителя чистоты науки (помните: не стать драконом - вот проблема!).
Наиболее эффективный метод борьбы с методологическим принуждением, как мне кажется, есть сознательное стремление каждого ученого к пониманию и владению не одной, а несколькими взаимоисключающими теориями (см. "Химию и жизнь" № 7 за 1988 г.). Когда понимаешь альтернативу, труднее быть фанатиком моноидеи, приобретаешь объемное видение проблемы.
Итак, пусть каждый из вас, прочтя эту статью, лично, наедине с самим собой, оценит меру своего методологического порабощения, способность противостоять ему, осваивая язык иного способа мышления.
ЧЕТЫРЕ ФУНДАМЕНТАЛЬНЫХ СВОЙСТВА КЛЕТКИ
Какую бы сторону жизнедеятельности клетки вы не объясняли, в ваши рассуждения так или иначе обязательно включены представления о том:
(1) почему одни вещества легко проникают в клетку, другие труднее, третьи практически в нее не проникают (свойство полупроницаемости);
(2) почему вещества, например, ионы K+ и Na+, распределяются между клеткой и средой неравномерно: одних больше в клетке, чем в среде, других - наоборот меньше (свойство избирательности);
(3) каким образом клетки генерируют электрические потенциалы (свойство генерировать разность потенциалов);
(4) каким образом клетке удается поддерживать осмотическое равновесие со средой (осмотические свойства).
В зависимости от того, как вы объясняете эти свойства, коренным образом зависит вся система ваших представлений о клетке.
ОСНОВНОЙ ВОПРОС ФИЗИОЛОГИИ КЛЕТКИ
Как это ни покажется странным, но основной вопрос состоит в том, в каком состоянии находится внутриклеточный K+: в свободном (растворенном в воде цитоплазмы) или связанном? В зависимости от варианта ответа на этот вопрос, вы будете вынуждены диаметрально противоположным образом выстраивать объяснения фундаментальных свойств клетки.
В самом деле, если K+ действительно свободен, то избирательность клетки к нему можно объяснить только особыми свойствами поверхностной мембраны - специально созданными в ней для калия каналами и насосами (закачивающими K+ внутрь клетки). Если же исходить из того, что K+ связан, тогда избирательность клетки к K+ и ее способность аккумулировать его в себе объясняется уже свойствами соответствующих центров связывания. При таком понимании нет очевидной необходимости в мембранном калиевом насосе.
Другие фундаментальные свойства также объясняются диаметрально противоположным образом в зависимости от ответа на основной вопрос. Так, если K+ связан, то рушится представление о трансмембранном потенциале, возникающим благодаря разности в концентрациях свободных K+ и Na+ по обе стороны поверхностной мембраны клетки: ведь свободным K+ остается только вне клетки. Осмотическая стабильность клетки, согласно традиционному подходу, также зиждется на свободе внутриклеточного K+ - основного противовеса осмотически активным компонентам среды. А если этой свободы нет - значит традиция несостоятельна.
В этой проблеме важно отметить, что историю экспериментальных исследований состояния K+ внутри клетки можно разделить на два периода - косвенных и прямых методов.
Работы первого периода (до 60-х годов), использовавшие в основном электрометрические методы, однозначно свидетельствуют о свободе клеточного K+: его активность и подвижность были такими же, как и в соответствующих водных растворах. Правда, были данные и о том, что активность и подвижность клеточного K+ могут почему-то значительно превышать таковые в водном растворе, но эти работы так и остались незамеченными, поскольку не поддавались логическому объяснению.
В последующий затем период, когда были привлечены более технологически сложные прямые методы (на основе микроскопической и электронно-микроскопической техники), воочию (в буквальном смысле) предстала связанность K+ с внутриклеточными структурами. Причем, исследования этого второго периода развивались в основном независимыми от представлений Линга исследователями.
ПРЕДСКАЗАТЕЛЬНАЯ СИЛА ТЕОРИИ, или А-ДИСКИ ПРОТИВ МЕМБРАН
Одна из центральных посылок теории Линга состоит в том, что клеточный K+ связан внутриклеточными структурами и вся физиология клетки должна строится именно на этом основании. Будучи катионом, он взаимодействует с фиксированными отрицательными зарядами, представляющими собой, главным образом, свободные карбоксильные группы остатков бикарбоновых аминокислот (аспарагиновой и глютаминовой), входящих в состав белков. Анализ показал, например, что 60% всех свободных карбоксильных групп поперечно-полосатой мышечной клетки, способных взаимодействовать с K+, расположены в А-дисках миофибрилл, основным компонентом которых является сократительный белок миозин. Следовательно, не менее 60% внутриклеточного K+ должно быть локализовано, согласно теории Линга, именно в миозиновых А-дисках.
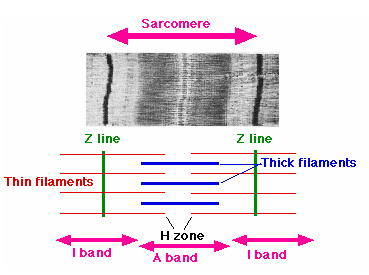
Дореволюционный подход дает прямо противоположное предсказание: поскольку внутриклеточный K+ свободен, т.е. растворен в воде, то калия должно быть больше в тех частях клетки, где больше воды, а воды больше в I-дисках миофибрилл [1], основным компонентом которых является другой сократительный белок - актин. Чтобы не быть обвиненным в том, что я сам приписываю K+ слишком большую свободу, играющую такую принципиальную роль, сошлюсь на сугубо научную литературу: "...Неорганические ионы внутри клетки находятся в основном в свободном состоянии и их накопление или выведение не может быть обусловлено избирательным связыванием ионов внутриклеточными структурами..." (П.Г. Костюк. Электрические явления на поверхностной мембране при активном транспорте ионов. В кн.: Структура и функции биологических мембран. М.: Наука, 1975, с.183-196). Еще одна выдержка: "Для мембранной теории биопотенциалов исключительно важное значение (выделено мной - В.М.) имело установление того факта, что большая часть ионов K+ в протоплазме находится в свободном состоянии, т.е. что протоплазма в соответствии с исходным предположением Бернштейна представляет собой свободный раствор этих ионов" (Ходоров Б.И. Общая физиология возбудимых мембран. М.: Наука, 1975, с. 8). Итак, кто же оказался прав? Где K+ больше - в А- или I-дисках?
Вопреки авторитетным заявлениям, революционные ожидания подтвердились самым блестящим образом. Методы тонкого внутриклеточного осаждения K+, авторадиографические методы с применением нестабильного изотопа K+, рентгеновский микроанализ (когда очень тонкий рентгеновский луч просвечивает отдельные части клетки), методы, основанные на выжигании лазерным лучом тех или иных клеточных структур, все эти методы прямо, без дополнительной интерпретации, свидетельствовали о том, что одним только миозином связывается не менее 60% внутриклеточного K+. Связанность K+ делает понятным и такой факт: при контакте с 85%-ым глицерином мышца в течение часа теряет 80% воды и только 10% (!) калия [2], и это при том, что соли калия (в частности, KCl) хорошо растворимы в глицерине. Приходится констатировать, что консервативный подход потерпел на этом участке фронта тяжкое поражение.
Таким образом, теория Линга удовлетворяет одному из требований научного метода - она способна давать принципиально важные предсказания, поддающиеся экспериментальной проверке. Кроме того, проверка проведена многократно, различными и независимыми группами исследователей, т.е. удовлетворено и другое ключевое требование научной методологии - экспериментальный результат воспроизводится в разных руках и в различных точках земного шара.
Как бы мы ни оценивали теорию Линга, факт локализации большей части K+ на миозиновых структурах мышечной клетки - самый твердый камень в ее основании.
ЭЛЕМЕНТАРНАЯ СТРУКТУРНАЯ ЯЧЕЙКА КЛЕТКИ
Согласно Лингу, элементарной структурной ячейкой клетки является ионо-водо-белковый комплекс. Хотя белок и упомянут последним, именно он является ядром комплекса и определяет все его основные свойства. Три структурные особенности белковой молекулы являются ключевыми: наличие свободных карбоксильных групп, необходимых для связывания катионов; геометрически правильное чередование пептидных связей, обладающих дипольным моментом; и центры связывания для так называемых управляющих лигандов: АТФ, Са2+, гормонов и пр.
Молекула белка линейной формы с геометрически правильным чередованием диполей пептидных связей (кислород - отрицательный диполь, азот - положительный) является идеальной матрицей, благодаря которой внутриклеточная вода структурируется. Дипольные заряды всех его пептидных связей, взаимодействуя с дипольными молекулами воды, ориентируют их в пространстве и ограничивают подвижность. Первый слой ориентированных и связанных белком молекул воды является, в свою очередь, матрицей для следующего водного слоя и т. д. В результате, вокруг белка образуется многослойная водная шуба. Чем больше доля пептидных связей данной белковой молекулы доступна воде (что зависит от конформационного состояния), тем эффективней такой белок структурирует воду. Рассмотрим некоторые свойства ионо-водо-белкового комплекса, постулируемые теорией Линга.
Управляющие лиганды обладают способностью изменять электронную плотность на различных функциональных группах белка (белок обретает черты сходства с молекулярным компьютером). При связывании, например, АТФ, электронная плотность на карбоксильных группах меняется таким образом, что у них повышается сродство к K+. K+ начинает аккумулироваться ионо-водо-белковыми комплексами клетки и в результате клетка в целом оказывается способной накапливать этот катион. Сам белок при этом "распрямляется", больше его пептидных связей начинают взаимодействовать с водой, образуя вокруг белковой молекулы более обширный водный кокон. Соседние коконы сливаются в единую систему и, как следствие, содержание связанной воды в клетке резко возрастает. В такой структурированной водной среде могут оставаться только молекулы и ионы достаточно малых размеров, которые легко вписываются в "кристаллическую" решетку многослойной системы ориентированных молекул воды. Na+ в эту систему вписывается плохо из-за слишком большой гидратной оболочки и потому покидает сначала объем, занятый ионо-водо-белковым комплексом, а затем, по мере сливания комплексов, и клетку.
Если АТФ расщепляется, то события приобретают противоположную направленность: K+ и вода сбрасываются, значительная часть пептидных связей белка уходит от контакта с водой, Na+ замещает K+ в центрах его связывания. Такие циклы связывания-расщепления АТФ и соответствующие этому изменения в структуре и свойствах ионо-водо-белкового комплекса (вместе с вкладами других управляющих лигандов) лежат, согласно Лингу, в основе функционирования отдельных белков, белковых комплексов, клеточных структур (в том числе мембран), органелл и клетки в целом.
ФУНДАМЕНТАЛЬНЫЕ СВОЙСТВА КЛЕТКИ С РЕВОЛЮЦИОННЫХ ПОЗИЦИЙ
Свойство № 1. Клетка полупроницаема не потому, что полупроницаема ее поверхностная мембрана с гипотетической системой отверстий разного калибра (теория сита), а потому что структурированная клеточная вода (где бы она ни была - и в мембране, и в цитоплазме) является хорошим растворителем для одних веществ (легко проникают в клетку) и плохим для других (плохо или совсем не проникают в клетку).
Свойство № 2. В клетке аккумулируются только те вещества, для которых в ней имеется достаточное количество центров связывания. Чем больше таких центров и чем выше их сродство (избирательность) к данному веществу (иону), тем больше разница между внутри- и внеклеточной концентрациями данного соединения (иона).
Свойство № 3. Электрический потенциал покоящейся клетки является адсорбционным потенциалом - результатом связывания ионов с белками микроскопически тонкого поверхностного слоя клетки, непосредственно контактирующего с внешней средой. Величина потенциала определяется сорбционными свойствами белков пограничного слоя: плотностью фиксированных зарядов, их избирательностью и сродством к противоионам.
Свойство № 4. Осмотическое равновесие клетки со средой поддерживается не за счет взаимодействия свободной внутриклеточной воды со свободным K+, а в результате связывания воды с белками. Осмотическая активность белка определяется величиной доли пептидных связей, доступной для взаимодействия с водой.
ЭНЕРГЕТИЧЕСКОЕ ИНАКОМЫСЛИЕ
Начиная с 50-х годов представление о роли АТФ, как универсального источника энергии для биологических процессов, не изменилось. Однако изменилась оценка того количества энергии, которое эта молекула способна дать: с 10-12 ккал/моль тогда, она снизилась до 3-5 ккал/моль в 70-е годы. Девальвация АТФ ставит вопрос об энергетическом крахе клетки, если смотреть на ее энергетику с консервативных позиций. Даже если верна первоначальная оценка калорийной ценности АТФ, то для обслуживания только натриевого насоса (учитывая реально наблюдаемую интенсивность обмена Na+ между клеткой и средой) необходимо в 30 раз больше АТФ, чем клетка способна синтезировать.
Линг полагает, что энергия, используемая клеткой для своих нужд, содержится не в отдельно взятой химической связи, а в целой системе взаимодействий в рамках АТФ-ионо-водо-белкового комплекса. При распаде этого комплекса энтропия системы возрастает (в основном за счет разупорядочивания ранее связанной и структурированной воды), что и служит источником свободной энергии для биологической работы. Для поддержания метастабильного состояния АТФ-ионо-водо-белкового комплекса не требуется непрерывного притока энергии. Энергетика клетки, по Лингу, чрезвычайно экономна.
ИМЕЮТ ЛИ ТЕОРИИ ПРАВО НА ИСКЛЮЧЕНИЯ?
Когда универсализм основных положений теории исчерпан и уже не может служить логической основой для объяснения всей совокупности явлений, появляются объяснения специальные, предназначенные только для данного, единичного, случая и непригодные для других частных случаев - гипотезы ad hoc. Ну, например, если утверждается, что различие в концентрации того или иного вещества внутри и вне клетки может поддерживаться только за счет деятельности специального мембранного насоса, то как быть с таким исключением, как кислород? В самом деле, с одной стороны существует универсальное правило: "Если вещество нейтрально, то вопрос о том, нужен ли насос в поверхностной мембране для перекачки данного вещества, решается очень просто: раз концентрация отличается, значит, нужен насос" (Е. А.Либерман. Живая клетка. М.: Наука, 1982, с.101). С другой стороны, концентрация кислорода в эритроците в 100 раз превышает его содержание в плазме крови, а каких либо кислородных насосов в мембране красной клетки нет. Универсализм нарушен!
В примере с кислородом всем нам повезло - доказано, что кислород связывается с гемоглобином. А если бы не было доказательств этому? Тогда бы не избежать нам кислородной помпы! А вот обратная ситуация: все, кто заглядывал в электронный микроскоп, знают, что большая часть K+ мышечной клетки связана с миозином. Тем не менее, калиевый насос продолжает существовать (видимо тем, кто работает с микроскопом, нет дела до насосов, а "насосники" не смотрят в микроскоп).
В полном соответствии с универсальным правилом мембранной теории, пришлось придумать множество разных насосов. Всякий раз, когда обнаруживается, что какое-то вещество распределяется между клеткой и средой неравномерно, сразу же постулируется специальная мембранная помпа, даже если клетка сталкивается с этим веществом впервые в своей непростой эволюционной судьбе. Уже насчитывается более 25 (!) разновидностей насосов (работа каждого из которых требует опять же непрерывного притока энергии) и этот список открыт для новых членов. Когда число систем активного транспорта перевалит за 1000, может быть тогда станет очевидной абсурдность такого способа мышления?
Но абсурдность видна и в другом. Вот один из примеров. Если лимфоциты человека охладить до 0 град. C в среде с нормальным соотношением K+ и Na+ (5 и 145 мМ соответственно), то концентрации Na+ внутри и вне клетки выравниваются. Как это объяснить с консервативных позиций? Довольно просто: натриевый насос, который постоянно требует для своей работы энергию в виде АТФ, перестает работать при 0 град.С, т.к. теплолюбивые лимфоциты при такой низкой температуре перестают синтезировать этот универсальный носитель энергии. Поскольку энергии становится недостаточно для эффективной работы насоса, концентрации Na+ по обе стороны мембраны выравниваются. Все гладко, не так ли?
Однако стоит уменьшить содержание Na+ в среде до 19 мМ (изоосмотичность среды поддерживалась при этом сахарозой), как наши лифоциты начинают вести себя весьма странно: при той же нулевой температуре в клетке начинают одновременно накапливаться и Na+, и K+: их становиться в клетке в 6 и 10 раз больше, чем в среде, соответственно [3].
Во-первых, получается, вопреки священному постулату, что натриевый насос "смонтирован" в мембране не жестко. На самом деле он может "качать" свой катион куда захочется ему, а не теории, его породившей.
Во-вторых, выходит, что Na,K-насос транспортирует свои катионы не в разные, как это всегда было, а в одну и ту же сторону по отношению к мембране (не слишком ли?). Наконец - энергия. Раз уж мы договорились, что при 0 град. С метаболизм прекращается, то тогда за счет какого источника энергии осуществляется поддержание пусть "уродливого", но градиента - когда и K+ и Na+ скапливаются по одну и ту же сторону мембраны?
Уже на одном этом примере мы сталкиваемся с впечатляющими противоречиями трем фундаментальным положениям консервативной парадигмы: монтаж насоса не жесткий, а произвольный; K+ и Na+ качаются в одну, а не в разные стороны; энергии нет, а градиент есть!
Есть и другие несуразности. Каждому, кто интересуется физиологией и биофизикой клетки, известно, как чувствительны клетки к осмотическому давлению среды: в гипотонических растворах клетки начинают набухать, в гипертонических - наоборот, их объем начинает уменьшаться, клетки съеживаются. Почему? В любом учебнике вы найдете объяснение, что всему причиной полупроницаемая мембрана, разделяющая два одинаковых по сути водных раствора: внутри- и внеклеточный. Но как тогда быть с тем, что мышечные волокна, лишенные мембраны все равно обладают осмочувствительностью [4]. А микросферы и родственные им коацерваты, прототипы древнейших одноклеточных организмов, не "нюхавшие" липидов и лишенные тем самым удовольствия иметь на своей поверхности бислойную липидную полупроницаемую мембрану? Эти безмембранные протоклетки осмочувствительны [5]! Значит, остается признать: наличие полупроницаемой мембраны не является критическим.
Очень некрасивый номер выкинули амебы: в эксперименте их лишили святая святых консервативной клеточной физиологии - все той же мембраны. Но несмотря на эту утрату и вопреки ей, они продолжали себе ползать как ни в чем ни бывало [6]. Мембраны нет, значит нет ни полупроницаемости, ни ионных насосов, ни биопотенциалов, ни особого внутриклеточного состава, но "раздетая" клетка, несмотря ни на что, продолжает демонстрировать нам одно из ярчайших проявлений жизни - движение. Так все-таки, нужна или нет поверхностная мембрана?
Сторонники общепринятых представлений так и не попытались дать последовательного, СОГЛАСОВАННОГО объяснения ВСЕХ основных вопросов клеточной физиологии. Нарастание числа гипотез ad hoc - свидетельство кризиса теории, ее неспособности на последовательную реализацию своих центральных идей в силу их несовершенства или ложности. Ну, а когда обнаруживаются явления и вовсе не поддающиеся как-либо трактовке, как в случае особо показательного примера с лимфоцитами, то на ум приходит горькая ирония Томаса Генри Гексли: "Трагедия науки: прекрасная гипотеза, убитая маленьким уродливым фактом".
Революционность Линга в том, что он сделал серьезную попытку предложить такие постулаты, которые, логически развертываясь, объединяют всю систему физиологических явлений в единую, стройную систему без грубых аномалий. Взгляды Линга являются попыткой создать подлинную теорию в классическом смысле, а не набор удобных частных правил, которые в любой момент разрешается менять на новые, еще более удобные.
КОНТРРЕВОЛЮЦИОННЫЕ ВОЗРАЖЕНИЯ
Возражение: не существует материалов, веществ, умершвленных препаратов живых клеток, изолированных белков, которые "предпочитали" бы связывать K+ в присутствии Na+. А раз так, то способность живой цитоплазмы избирательно связывать K+, проповедуемая Лингом, является надуманной, не поддающейся проверке.
Ответ: одной из самых известных систем, способных избирательно связывать K+ является обыкновенная почва. Некоторые минералы (пермутиты), ряд синтетических ионо-обменных смол, обладают повышенным сродством к K+. Специальные сорта стекол селективно связывают K+ в присутствии Na+. А в 1980 году Л. Эдельманн [7] впервые получил мертвый препарат мышечной клетки, который оказался способным избирательно связывать K+ в присутствии Na+. Все предыдущие попытки зарегистрировать сродство к K+ изолированных мышечных белков, глицеринизированных мышц оканчивались неудачей. Вероятно, только Эдельманну удалось сохранить в препарате мышцы некий фактор, без которого сродство к K+ сократительных белков утрачивается.
Возражение: каналы в мембранах видели в электронный микроскоп.
Ответ: да, структуры, называемые каналами, действительно существуют и играют важную роль. Но являются ли они каналами в обычном смысле слова, т.е. соединяющими два водных бассейна - внутриклеточный и внеклеточный? Внутриклеточная среда совсем непохожа на внеклеточную. Вне клетки - раствор, внутри клетки - сложная, организованная "коллоидная" система, свойства воды в которой существенно отличаются от свойств воды внеклеточной. С этим, пожалуй, сегодня согласится каждый, кто хоть и не читал Линга, но знаком с проблемой состояния воды в биосистемах. Таким образом, "канал" - просто неудачный, устаревший термин, т. е. это проблема не теории Линга, а чисто терминологическая проблема.
Возражение: хорошо известно, что тени эритроцитов, т.е. "выпотрошенные" эритроциты, без цитоплазмы, от которых осталась лишь их поверхностная мембрана, способны аккумулировать K+ и откачивать из полости "мыльного пузыря" Na+. Не доказывает ли это реальность Na,K-насоса?
Ответ: в специальных опытах Лингом было показано, что чем больше белков цитоплазмы содержится в "мыльном пузыре" в качестве примеси к мембране, тем успешнее он аккумулирует K+ и тем меньше в нем содержание Na+. Если тень эритроцита действительно не содержит примеси цитоплазматических белков, т.е. действительно является тенью, то она имеет самое призрачное отношение к проблеме перекачки катионов.
Однако гораздо удобнее работать с тенью аксона - гигантским аксоном кальмара, из которого удалена аксоплазма. Из-за своих больших размеров этот препарат не требует применения сложных технических ухищрений и процедур, с ним можно экспериментировать даже в школьном кабинете биологии. С появлением в 1961 году этой гигантской тени, мембранной теории, впервые за всю историю ее существования, представилась возможность поставить РЕШАЮЩИЙ ЭКСПЕРИМЕНТ для доказательства реальности Na,K-насоса: заменить аксоплазму морской водой ("родной" для аксона кальмара стихией) или другим раствором с преобладанием Na+ над K+, добавить в нее необходимые для работы "насоса" энергоносители (АТФ, аргининфосфат и т.п.) и посмотреть, будет ли Na+ откачиваться в окружающую среду, а K+ накапливаться внутри аксона-призрака с тем, чтобы со временем состав внутриаксональной жидкости приобрел типичный для живой клетки ионный состав (т.е. с преобладанием K+ над Na+).
В 1963 году известный электрофизиолог Кейнес (Keynes R.D.) сообщил на семинаре в Пеннсильванском университете, что попытки такого рода делались, но все они закончились неудачей. С тех пор решающий эксперимент на аксоне кальмара остается несбыточной мечтой мембранной теории.
Возражение: Но этот же пример с тенью аксона ясно показывает, что потенциал покоя и действия существует в неизменном виде и при отсутствующей аксоплазме. Более того, оболочка аксона заполнена разбавленным водным раствором электролитов, т.е. K+ в нем безусловно свободен, но это кардинальное, с позиций теории Линга, событие - освобождение K+, - ничего не прибавляет и не отнимает от свойств аксона.
Ответ: С точки зрения теории Линга, в этом возражении нет ничего парадоксального и тем более убийственного: за потенциал покоя и действия ответственна не аксоплазма, а лишь внешняя поверхность аксона, которая остается на своем месте и после превращения аксона в тень.
Как уже говорилось, потенциалы покоя и действия, по Лингу, являются адсорбционными. В случае с перфузируемым аксоном, регистрируемый потенциал является алгебраической суммой адсорбционных потенциалов на двух поверхностях аксолеммы: на наружной и внутренней, созданной искусственно.
Как и в случае с Na, K-насосом, тень аксона оказалась идеальным объектом для проверки мембранной теории биопотенциалов Ходжкина-Каца-Гольдмана (Hodgkin-Katz-Goldman), впервые опубликованной в 1949 году. Согласно уравнению, предложенному этими авторами, потенциал покоя обязан зависеть и от внутриклеточной концентрации K+ и Na+. Однако известный ученый Тасаки с сотрудниками [8-10] показали, что при широкой вариации концентраций K+ (от 150 до 500 мМ) и Na+ (от 0 до 350 мМ) в перфузируемой жидкости, "мембранный" потенциал практически не менялся! С позиций теории Линга этот результат объясняется просто: искусственная, внутренняя, поверхность аксолеммы не предназначалась природой для формирования адсорбционного потенциала - на ней нет связывающих центров в необходимом количестве и нужного качества. Поэтому он там практически отсутствует. То есть алгебраическая сумма потенциалов определяется в этой системе только потенциалом наружного слоя аксолеммы.
Возражение: чем не устраивает Линга теория сита? В липидном барьере мембраны существуют отверстия разной величины, в соответствии с которыми одни молекулы или ионы могут проникнуть в клетку, другие нет. Все просто и понятно!
Ответ: Простота обманчива и роковая роль теории сита для физиологии тому пример - до сих пор (!) она является отправной точкой для понимания полупроницаемости биологических мембран.
Автором теории является Мориц Траубе (Moritz Traube), изучавший проницаемость для различных веществ тонких пластин (мембран) из медно-ферроцианидного геля (образующегося при сливании растворов сульфата меди с ферроцианидом калия - опыт, доступный уже школьному кабинету химии) и первым обнаруживший явление полупроницаемости искусственных мембран. В 1867 г. он объяснил его существованием в мембране отверстий соответствующих размеров, благодаря которым она виделась (и видится до сих пор) атомным/молекулярным ситом.
Идея Траубе явилась толчком для ботаника Вильгельма Пфеффера (Wilhelm F. Pfeffer), который в 1877 году закладывает фундамент мембранной теории следующим предположением: протоплазма клетки окружена полупроницаемой, по аналогии с медно-ферроцианидной, мембраной. А в 1899 году Чарлз Овертон (Charles E. Overton), троюродный брат Чарлза Дарвина, уточняет: раз вещества тем легче проникают в клетку, чем они лучше растворяются в оливковом масле, значит мембрана Пфеффера, служащая барьером для диффузии веществ - липидная (о порочности этой логики см. [14]). С тех времен прочно и, похоже, навсегда закрепилось представление о том, что липидная мембрана - сито, разделяющее два водных раствора - внутри- и внеклеточный. В дальнейшем эта схема только уточнялась и по-своему развивалась, но никогда не подвергалась ни малейшему сомнению.
К 1937 году прошло так много лет со времен Траубе-Пфеффера-Овертона, что уже никто не обратил внимание на одно неприятное событие: методами дифракции рентгеновских лучей и электронов было установлено, что средний размер пор той самой медно-ферроцианидной мембраны во много раз превосходит размер молекул, для которых она была и остается непроницаемой [11].
Было экспериментально опровергнуто одно из ключевых положений мембранной теории - теория сита. Но что такое маленький фактик в сравнении с великой теорией?
Возражение: Речь шла о липидных мембранах. Здесь невозможно не вспомнить о валиномицине, соединении, способном избирательно связывать K+ на одной стороне искусственной липидной мембраны и освобождать его на другой стороне. Работая как переносчик, валиномицин будет "трудиться" до тех пор пока концентрация K+ по обе стороны мембраны не выровняется. Точно так же валиномицин действует и на живые клетки, например, на эритроциты: под его действием клетки теряют K+. Разве это сходство не доказывает многого: что липидная мембрана клетки действительно является барьером для K+, что внутриклеточный K+ свободен и что существование переносчиков в мембране клеток - реальность?
Ответ: "Доказательность" эффектов валиномицина - издержки методологической эйфории. Со времен Овертона утвердилось правило, превратившееся в бездумную традицию: если вещество гидрофобно (т.е. жирорастворимо), как в случае с валиномицином, то его биологическая активность объясняется тем, что оно растворяется в липидном (жирном) слое мембраны. Зависимость эффективности биологического действия химических агентов от их гидрофобности, открытая Овертоном, однозначно интерпретировали как результат агент-мембранных взаимодействий.
В традиции связывать всякое гидрофобное взаимодействие только с липидами мембраны была своя логика: нужно помнить о том, что на протяжении многих десятилетий науке была известна только одна гидрофобная область клетки - липидная фаза ее поверхностной мембраны, а затем и мембран других локализаций.
Однако сейчас логика умозаключений Овертона представляется полностью несостоятельной. С 60-х годов стало ясно, что белки также имеют гидрофобные области, ничем принципиально не отличающиеся по своей физической природе от гидрофобной фазы липидов. Поэтому, когда действие валиномицина на клетку безапелляционно связывают в наше время только с его встраиванием в липидный слой мембраны, это являет собой пример архаичного мышления. Совершенно ясно, что попадая в клетку, весьма и весьма гидрофобный валиномицин взаимодействует со всем, что есть в ней гидрофобного: и с липидами, и с белками, и белковыми комплексами, имеющими межбелковые гидрофобные области. В результате такого взаимодействия с клеточными белками (масса которых на порядки превосходит массу липидов), K+-связывающий матрикс клетки теряет способность удерживать K+; катион переходит в свободное состояние и, подчиняясь законам диффузии, выходит из клетки. Вот объяснение действия валиномицина на клетку, которое обходится без "обязательного", освященного вековой традицией, взаимодействия этого соединения с липидными барьерами [14].
Что же касается универсализма барьерной функции липидов, на которой, собственно, и держится вся мембранная теория, то небезынтересно будет вспомнить о внутренней мембране митохондрий, широко известной в узких кругах, которая, как оказалось, содержит так мало липидов, что уже "официально" отказались представлять ее как сплошной билипидный слой с вкраплениями каналов, переносчиков, насосов и пр. Наоборот, выяснилось, что эта мембрана представляет собой сплошную белковую структуру с редкими включениями липидов [12]. То есть, иначе говоря, Великую Барьерную Функцию Липидов в отдельно взятой митохондрии тихо отменили. Почему ее нельзя отменить и во всех остальных случаях?
ИСТОКИ РЕВОЛЮЦИИ
Столкновения взглядов на природу физико-химической организации клетки начинаются почти одновременно с формированием клеточной теории в 30-х годах XIX века. Первое заявление на этот счет сделал Феликс Дужардин (Felix Dujardin, француз), первооткрыватель протоплазмы, названной Томасом Хаксли (Thomas Huxley, 1825-1895) "физической основой жизни": он видел в ней желеподобную субстанцию, а не обычный раствор. И для этого были веские основания: уже первые наблюдатели отмечали, что протоплазма, выдавленная из растительной клетки, не смешивается с водой и сохраняет целостность.
Томас Грехем (Thomas Graham), посвятивший всю жизнь исследованию диффузии, делает в 1861 г. следующий шаг, имевший принципиальное значение для развития альтернатив мембранной теории - вводит понятие коллоидного состояния вещества и приходит к выводу, что вода активно взаимодействует с веществами биологического происхождения, например, с желатиной, крахмалом и т.п. При этом она переходит в связанное состояние и значительно меняет свои свойства как растворитель по сравнению с обычной водой. Именно благодаря связанной воде, как он полагал, тонкие пластины, приготовленные из различных гелей, обладали свойством полупроницаемости.
Далее Отто Бючли (Otto Bьtscli) был первым, кто в 1894 году высказал мысль, впоследствии надолго забытую общепринятыми представлениями, о том, что протоплазма является структурированной, организованной системой, а не гомогенным коллоидом и тем более не раствором. Для сохранения целостности такой протоплазмы, как он полагал, мембрана не требуется.
В 1905 году Макаллум (Macallum), используя метод внутриклеточного осаждения K+ кобольтонитритом натрия (sodium coboltinitrite) впервые показывает, что K+ локализован главным образом в А-дисках миофибрилл мышечных клеток.
В 1906 году Мур (Moore), Роаф (Roaf) и др. находят новые экспериментальные свидетельства связанного состояния внутриклеточного K+ и проводят аналогию между механизмами накопления клеткой K+ и кислорода эритроцитом.
Начиная с 1907 и по 1938 год выходит серия работ Мартина Фишера (Martin H. Fischer) и его сотрудников, в которых обосновывается взгляд на протоплазму как на "соединение белка, соли и воды в гигантскую молекулу".
1940 году выходит в свет монография Д.Н. Насонова и В.Я. Александрова "Реакция живого вещества на внешние воздействия", в которой впервые за многие годы давалась систематическая развернутая критика мембранной теории. Выдающийся цитолог, первооткрыватель функции аппарата Гольджи, Д.Н. Насонов разработал оригинальные цитологические методы, среди которых центральное место занимал метод исследования распределения между клеткой и средой органических анионов и катионов. Д.Н. Насонов пришел к выводу, что изменения в поглощении органических ионов покоящейся, возбужденной и поврежденной клеткой объясняются не изменением проницаемости клеточной мембраны, а связыванием этих ионов белками протоплазмы.
Учеником Д.Н. Насонова, А.С. Трошиным, была создана количественная сорбционная теория распределения между клеткой и средой неорганических ионов и нейтральных молекул.
Д.Н. Насонов рассматривал протоплазму как коллоидную фазу, несмешивающуюся с водой, обладающую особыми свойствами (поэтому это направление получило название фазовой теории) и с этих позиций дал последовательное и логичное объяснение многих проблем клеточной физиологии (см. монографию "Местная реакция протоплазмы и распространяющееся возбуждение", М.-Л., Изд-во Академии наук СССР, 1959).
Будучи последовательным, он отрицал значение и даже само существование клеточной мембраны. Поэтому легко понять, каким ударом для него явились присланные ему первые электронно-микроскопические фотографии биологических мембран с хорошо всем знакомой теперь трехслойной структурой. Несомненно, что такой поворот событий подорвал престиж фазовой теории в глазах обывателей от науки. Однако А.С. Трошин, сменивший Насонова на посту директора Института цитологии АН СССР, сохранил верность своим взглядам и систематизировал их в своей итоговой монографии "Распределение веществ между клеткой и средой" (Л.: Наука, 1985).
Однако время показало, что электронные фотографии мембран кажутся убийственными лишь на первый взгляд. Неожиданно выяснилось, что поверхность протеиноидных микросфер Фокса, которые рассматриваются как один из вариантов первобытной клетки, тоже дают типичную для липидных мембран трехслойную структуру, хотя не содержат в своем составе "ни грамма" липидов. Это доказывает, что поверхностные слои микросфер, коацерватов и клеток не обязательно должны содержать липиды, чтобы выглядеть "как надо". Об этом говорят и электронные микрофотографии мембран митохондрий, из которых экстрагировали липиды: на них видна все та же неизменная трехслойная структура [13].
Из сказанного следует, что липиды биомембранам несомненно нужны, но не для того, для чего они нужны мембранной теории.
Вернемся однако к ходу истории. Первые положения современной фазовой теории были высказаны Лингом в 1952 г. 10 лет спустя он издает обширную монографию "Физическая теория жизни: гипотеза ассоциации-индукции" (A physical theory of the living state: the association-induction hypothesis. Waltham (Massachusttes): Blaisdell, 1962), в которой детально рассматривает все "за" и "против" фазовых представлений и дает объяснение со своих позиций основных феноменов клеточной физиологии. Последующие 20 лет исследований были подытожены в следующей монографии "В поисках физической основы жизни" (In search of the physical basis of life. New York: Plenum Press, 1984). В ней мы находим фазовые теории электрических потенциалов, окислительного фосфорилирования, мышечного сокращения, активного транспорта через эпителии, подходы к механизмам синтеза белка, роста и дифференциации, канцерогенеза. Дальнейшее развитие этих идей представлено в последующих монографиях Линга (A Revolution in the Physiology of the Living Cell. Krieger Publishing Company, Malabar, Florida, 1992; Life at the Cell and Below-Cell Level. The Hidden History of a Fundamental Revolution in Biology. Pacific Press, 2001).
ЖИВИТЕЛЬНАЯ СИЛА НАУКИ
Фанатичная преданность какой-либо одной идее на самом деле тормозит развитие науки, а часто и общества. Фанатик везде и во всем видит подтверждение своей правоты. Но методологическое поощрение любимой точки зрения антинаучно, ибо ясно, что каждая теория допускает проверку самое себя только на своих собственных условиях и поэтому оказывается наилучшим образом защищенной от опровержений. Очень удобная и потому общепринятая позиция, получившая широчайшее признание добропорядочной науки.
Но я утверждаю: без постоянного насилия над добропорядочностью невозможны ни открытия, ни прогресс. Ведь понятно: была бы гипотеза, а нужные факты найдутся. Поэтому в научный обиход необходимо постоянно внедрять высшую форму противоречий - антагонизм идей и принципов, несовместимых с хорошо обоснованными теориями или фактами. Только при таких условиях будут рождаться новые данные, подвергающие необходимым и полезным испытаниям общепринятую точку зрения. Идеал ученого - специалист, владеющий логикой всех конкурирующих теорий, а не только любимой [15].
Дефекты теории обнаруживаются благодаря контрасту с ее антиподом, а не анализу. Идеи нужно сравнивать с другими идеями, а не с "опытом". Но те из них, которые терпят поражение в соревновании необходимо усовершенствовать, а не отбрасывать. "Делать слабое более сильным", как говорили софисты, и благодаря этому поддерживать движение целого. Ни одна из теорий не должна превращаться в жесткую хорошо оплачиваемую идеологию.
Наша задача - сохранить не старую, а лучшую теорию.
ЛИТЕРАТУРА
1. Huxley A.F. and Niedergerke R. Measurement of the striations of isolated muscle fibers with the interference microscope. J. Physiology., 144: 403-425, 1958.
2. Hummel Z. The effect of different alkali metal ions on the release of muscle potassium during glycerol treatment. Physiol. Chem. Phys. Med NMR, 15(2): 177-180, 1983.
3. Negendank W. and Shaller C. Simultaneous net accumulation of both K+ and Na+ by lymphocytes at 0o C. Bioc. Biop. Acta, 640: 368-373, 1981.
4. Elliott G.F. Donnan and osmotic effects in muscle fibres without membranes. J. Mechanochem. Cell Motil. 2: 83-89, 1973.
5. Фокс С. и Дозе К. Молекулярная эволюция и возникновение жизни. М.: Мир, 1975, с.220.
6. Pollard T.D. and Ito S. Cytoplasmic filaments of Amoeba proteus. I. The role of filaments in consistency changes and movement. J. Cell. Biol., 46: 267-289, 1964.
7. Edelmann L. Preferential localized uptake of K+ and Cs+ over Na+ in the A-band of freez-dried embedded muscle section: detection by X-ray microanalysis and laser microprobe mass analysis. Physiol. Chem. Phys., 12: 509-514, 1980.
8. Tasaki I. and Takenaka T. Resting and action potential of squid giant axons intracellulary perfused with sodium-rich solutions. Proc Natl Acad Sci USA 50: 619-626, 1963.
9. Tasaki I. and Takenaka T. Eeffects of various potassium salts and proteases upon excitability of intracellularly perbused squid giant axons. Proc Natl Acad Sci USA 52: 804-810, 1964.
10. Tasaki I., Luxoro M., and Ruarte A. Electrophysiological studies of chilean squid axons under internal perfusion with sodium-rich media. Science 150(698): 899-901, 1965.
11. Fordham S. and Tyson J.T. J. Chem. Soc. 1937: 483, 1937. Обзор других работ такого рода см. обзор Glasstone S. Textbook of Physical Chamistry, 2nd ed., New York: Van Nostrand, 1946.
12. Sjostrand, F.S. The structure of mitochondrial membranes: a new concept. J Ultrastruct Res. 64(3): 217-245, 1978
13. Fleischer S., Fleischer B., Stoeckenius W. Fine structure of lipid-depleted mitochondria. J Cell Biol. 32(1): 193-208, 1967.
14. Matveev V.V. Protoreaction of protoplasm. Cell Mol Biol. 51(8): 715-23, 2005.
15. Matveev V.V., Wheatley D.N. "Fathers" and "sons" of theories in cell physiology: the membrane theory. Cell Mol Biol. 51(8): 797-801, 2005.
Подготовлена с использованием материалов книги Гильберта Линга "Физическая теория живой клетки"
В.В. Матвеев, Революция в физиологии клетки // «Академия Тринитаризма», М.,
|
|