|
|
|
Сергей А. Алферов
Новая публикация Бориса Розина заставила еще раз пройти по указанным в ней ссылкам (в частности и к источнику на). Вызвала восхищение гипотеза Б.Розина![]() 1
1
Автор использует понятие «асинхронности», то есть неодновременности действия, попеременности включения элементов в общем цикле. В нашем случае «асинхронность» — это не одинаковая включенность клеток в действие после их появления, это размножение не всех клеток сразу, а поочередно, с паузами.
В тексте «О Родительском ряде» наличие паузы «становления» перед включением в общее развитие явилось важным отличием процесса «Родительского ряда» от «механистичных процессов» наподобие цепной реакции. Теперь эти темы соединяются.
Приведем формулировку автора: «При асинхронном делении каждая клетка делится на две клетки, одна из которых пропускает следующий такт деления. …Асинхронность есть выражение асимметричности во времени.»
Сформулируем гипотезу Б.Розина в таком виде.
В каждый определенный момент (в практической фиксации этого момента![]() 2
2
Сделаем следующую схему процесса деления зиготы.
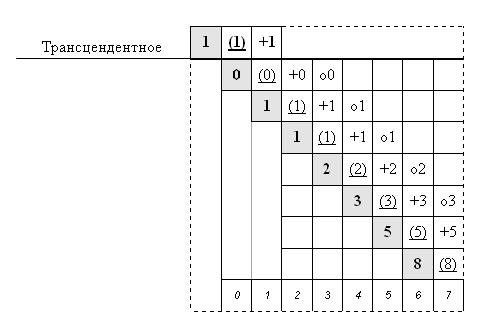
Ниже прокомментируем использованные обозначения.
|
Содержание i-того этапа деления |
|
|
оN1 |
«Отдыхающие» клетки после этапа
(i-1)
(Входят в количество «исходных») |
|
+N2 |
Редуплицированные из «исходных клеток» этапа
(i-1)
(N1 + N2 = N3) |
|
(N3) |
«Исходные клетки» этапа «i»
(«результирующие i-1»)
(N2 + N3 = N4) |
|
N4 |
Результирующее количество клеток этапа «i» |
Соответственно 4 элемента каждой строчки будут означать по порядку:
1. n – результирующее количество на этапе (i)
2. (n) – исходное количество на этапе (i+1)
3. +n – редуплицированное количество на этапе (i+2)
4. оn – «отдыхающее» количество на этапе (i+3)
Здесь есть интересный вопрос. Клетка, делясь на две, воспроизводит (клонирует) две одинаковые. Когда происходит изменение в клетке для дифференциации организма (тканей)? Понятно, что это может быть только в период «паузы деления»…
Но главным здесь пока остается вопрос подтверждения гипотезы. Борис Розин говорит, биологи «теперь знают, что искать», но видимо подтверждения пока нет… А давайте спросим, могут ли быть другие варианты асинхронного (по частям) размножения редуплицирующихся клеток?
Другого варианта, как деления клетки на две, у нас нет. Значит, мы можем только в момент выделения новой группы удвоения использовать какой-то иной алгоритм ее получения. То есть выделить на следующий этап какие-то другие части «отдыхающей» и «размножающейся» групп. Причем пропорция такого разделения должна быть всегда одинаковой.
И вот здесь – всё! Во-первых, мы не можем придумать такие части (делители), которые всегда обязательно смогут быть одинаковы. Здесь могут быть только 2 варианта: или размножаться целиком обеим группам (и тогда это синхронный вариант), или «отдыхавшие» берутся целиком плюс половина от «удвоившихся» (что и есть). А, во-вторых, «не надо умножать сущности», всё уже четко, элегантно, взаимообусловлено. Это значит, процесс естественного размножения из одного элемента: асинхронный и удвоением, – может происходить поэтапно только числами Фибоначчи.
Модель асинхронного морфогенеза клетки может быть только такой. Другое дело, что в реальности процесс деления может накладываться какими-то фазами соседних этапов. Нужны критерии и эксперименты, окончательно определиться в этом вопросе, как говорит Б.Розин, должна практика… Хотя, судя по информации из «Акушерства», они уже определились для начального этапа деления. А дальше и не обязательно; очевидно, что алгоритм « деления/умножения клеток вдвое только того количества, которое было всего на предыдущем этапе» должен сохраниться. Правда, вопрос – как из только что разделившихся клеток половина отбирается для «отдыха» при второй идентичной (!?) половине снова включающейся в деление (вместе с предыдущими отдохнувшими)? Кто/что запускает такой механизм и поддерживает его?..
Деление клеток зиготы по числам Фибоначчи — это не задача о кроликах, которая, во-первых, придуманная, а во-вторых, конкретное решение которой зависит от условий. Вот теперь видно, что по своей родовой сути – это действительно «Родительский ряд».
Родительский ряд создается самым простым и единым (универсальным) алгоритмом попеременного удвоения частей целого. И нет другого универсального (на всем протяжении ряда) алгоритма получения нового члена через редупликацию части предыдущего члена. Формальная запись этого свойства тривиальна: Fn+1 = 2ґ Fn-1 + Fn-2, — но дело не в том, что она выводится из канонической записи для члена ряда Фибоначчи. Дело в том смысле, который скрывается за этой записью.
Эти смыслы аддитивного ряда настолько важны и настолько являются сущностными для самого ряда, что родовое именование «Родительский ряд» перевешивает значение Фибоначчи, намного перевешивает значение привезенной от арабов «задачи о кроликах» и значение гораздо позже присвоенного именования. Новое именование также заслуживает использования.
Какие здесь есть еще общие выводы, вопросы, сентенции?
1. В первой «структурной таблице» можно заметить:
- элементы одного количества в горизонтали показывают изменение качества (состояния),
- в теле «количеств» легко выделяются квадры (без «отдыхающих), что может помочь исследовать ее отношения,
- существует вопрос отношений «трансцендентного» и «материального», проявляющийся в том числе в «самопорождении»; верхняя левая часть является некой иллюстрацией этого…
2. Новое качество созревает в паузе. Пауза порождает развитие.
3. И конечно, хочется еще раз заметить соответствие нижних структур.
Левая структура идет по одной всегда размножающейся веточке (правой) правой структуры.
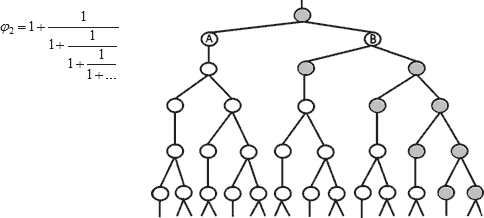
4. И может быть в этой ветвящейся структуре есть подсказка для представления в ветвящихся цепных дробях оставшихся обобщенных чисел ряда ЗП (1,465, 1,380 и т.д.)?...
* * *
Вроде бы тема пока закончена… Но вот эти упомянутые варианты задачи о кроликах !… Что это за варианты? По условиям задачи, взрослея, пара кроликов пропускала один первый такт размножения. А если пауза больше: 2 или 3 такта?... Этот вопрос аналогичен и для клеток, ведь кролики, размножаясь, тоже удваиваются (по условиям задачи).
Итак, вот схема нового процесса. И справа те суммарные количества, которые при этом образуются.
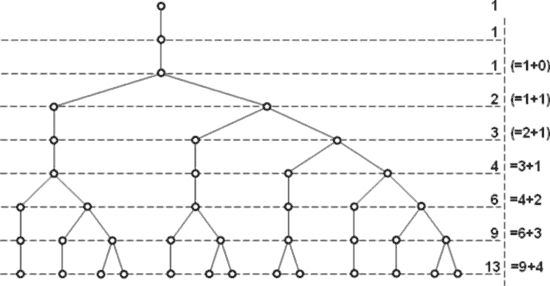
Можно нарисовать аналогичную схему для паузы, равной 3-м тактам. Тогда новые суммарные количества образуют ряд:
1 – 1 – 1 – 1 – 2 – 3 – 4 – 5 – 7 – 10 – 14 – 19 – 26 – ….
Видно, что это аддитивные ряды с суммированием членов через какое-то их количество. Первый ряд — через 1 член, следующий — через 2 …
Первый раз о «математическом существовании» таких рядов я узнал от ![]() 3
3
Итак, каждый ряд формировался суммированием предыдущих членов через свое порядковое их количество. Ряд Фибоначчи, дающий «1,618», — через 0 членов;
ряд, дающий «1,465», — через 1 член;
ряд, дающий «1,380», — через 2 члена;
ряд, дающий «1,324», — через 3 члена;
ряд, дающий «1,285», — через 4 члена;
и так далее. Очень красиво…
Приближение к «заданному индексу» происходило «волнообразно», то сверху, то снизу в некоторой группе значений.
Значения начинали суммироваться, когда количество первых членов становилось больше «шага суммирования», а первыми членами были выбраны значения классического «родительского ряда». Мне представилось, что этот ряд нужно начинать именно единицами до момента, когда можно будет сделать первое интервальное сложение. Теперь можно утверждать, что это именно так. Количество единиц при этом равняется величине (шаг+2) или значению «n» (см. дальше).
Итак, мы имеем новую структурную схему, в которой находится тот ряд, что дает отношением своих соседних членов очередное после «1,618» число «1,465». И точно также она может подсказать вид цепной дроби, дающей это число. Ведь до сих пор эта задача не решена ни для одного числа, кроме j 2=1,618...
Как эта структурная веточка «паузы» должна повлиять на структуру цепной дроби? Какой структурой цепной дроби изобразить переход на следующий уровень без деления? Да еще такой, которая будет иметь общую логику изменения в зависимости от количества «пауз»…
Например, так: 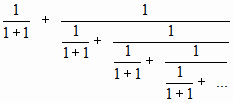
Значение этой дроби равно 1,280776… А если будет 3 «единицы» в знаменателе, то «1,180460…». И вот здесь настало время вспомнить об одной таблице.
В тексте «О взаимообратных числах» была приведена следующая таблица, в основу которой легли вот эти 2 известных уравнения:
и формула для варианта «обобщения ЗП»: 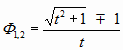
|
t |
Взаимообратные | n | |
| x (Ф1) | y (Ф2) | ||
|
0 |
0 |
Ґ |
ѕ |
|
0,5 |
j 13 |
j 23 |
0,1865266 |
|
1 |
Ц 2- 1 |
Ц 2+1 |
0,6067801 |
|
11/3 |
0,5 |
2 |
1 |
|
2 |
j 1 |
j 2 |
2 |
|
2,5534846 |
0,6823278 |
1,4655712 |
3 |
|
3 |
0,7207592 |
1,3874258 |
3,8958017 |
|
3,0497772 |
0,7244919 |
1,3802776 |
4 |
|
3,5097554 |
0,7548777 |
1,3247179 |
5 |
|
3,9439219 |
0,7780896 |
1,2851990 |
6 |
|
4 |
0,7807764 |
1,2807764 |
6,1328032 |
|
4,3584517 |
0,7965443 |
1,2554229 |
7 |
|
4,7573478 |
0,8116523 |
1,2320546 |
8 |
|
5 |
0,8198039 |
1,2198039 |
8,6250372 |
|
5,1433825 |
0,8243006 |
1,2131497 |
9 |
|
5,5185751 |
0,8350790 |
1,1974914 |
10 |
|
5,8844537 |
0,8443976 |
1,1842762 |
11 |
|
6 |
0,8471271 |
1,1804604 |
11,3206560 |
|
7 |
0,8672954 |
1,1530097 |
14,1852191 |
|
8 |
0,8827822 |
1,1327822 |
17,1942383 |
Величина «2/t» равна соответствующей разности «Ф2-Ф1». Можно взять любое число, вычислить обратное ему, а потом их разность – это будет отношение «2» к общему их оператору «t».
На системный характер этого оператора указывало также следующее обстоятельство. В виде простой цепной дроби (в том числе с начальным общим слагаемым) можно выразить любое число; в этом случае это слагаемое образуется разностью этого числа с его обратным числом:
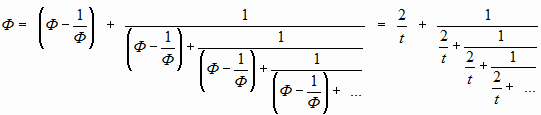
Мы сделали «вводную». А теперь посмотрим таблицу. При 2-х «единицах» в знаменателе мы получили значение Ф2 при t=4. При 3-х «единицах» — Ф2 при t=6… А ещё интересно посмотреть на правую часть верхней формулы. То есть наш «многоединичный знаменатель» — это «t/2»…
Тогда понятно, что при значении знаменателя (1+0,5) эта дробь дает табличное значение для «t=3». А при (1+0,7548777) – «n=5». (Заметьте красивую странность равенства последнего нецелого числа и значения в строке n=5.)
Итак, для структуры с «единицами» в знаменателе имеем:
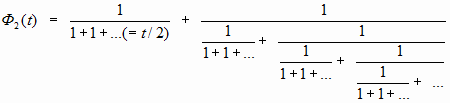
Но решения задачи представления числа ОЗС избранная структура цепной дроби не дает. Более того, вроде бы подсказывает, что нет решения на этом пути. Всё дело в той «странности». Левая (малая) дробь должна быть конечной (в пока существующем понимании соответствия структур). Если для «странного» варианта пытаться как-то структурно продолжать эту дробь, то потребуется искать представление для 0,7548777…=1/1,324717… А последнее число в знаменателе не имеет конечной дроби. В нашем же представлении она должна быть конечной. Если не получается этот вариант, то не подходит и всё представление…
Не дает продолжения и такое красивое решение:
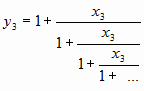
Итак, решение ОЗС в цепных дробях мы не нашли. Но оно очень близко. Очень…
А как же другие варианты задачи о кроликах? Например, когда пара приносит не одну пару, а скажем, 2, или 3, или 4?... Как и бывает в жизни; размножение кроликов – не морфогенез клеток… Конечно, это будет какой-то новый ряд… И структурная схема будет более ветвящейся. И по ней можно сделать структуру соответствующей ветвящейся цепной дроби… Но что-то я не верю, что там находятся искомые нами…
 1)
1)- Профаза – четкое структурное формирование хромосом с удвоенным кодоном.
- Метафаза – формирование веретена (перегородки) в ядре.
- Анафаза – разделение хромосом на две и расхождение к полюсам.
- Телофаза – разделение ядра и клетки.
Сергей А. Алферов, Родительский ряд и морфогенез // «Академия Тринитаризма», М.,
|
|