|
|
|
П.П.Гаряев, Валтрауд Вагнер, Е.А.Леонова-Гаряева, А.Е.Акимов, И.А.Сухоплюева, Э.Р.Мулдашев
В этой работе мы показываем, что воздействие на препарат ДНК in vitro определенными сочетаниями электромагнитных полей в диапазоне УФ-ИК вызывает ответ в форме многообразных, динамичных, долгоживущих реплицирующихся волновых отображений ДНК и ее ближайшего окружения.
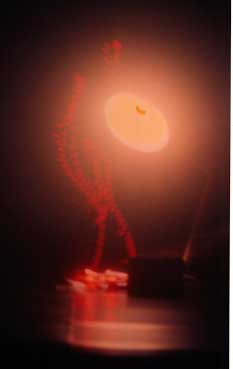
Мы обнаружили способность препаратов ДНК in vitro давать многократно реплицирующиеся отклики-отображения, реплики самой ДНК и ее ближайшего окружения. Такие реплики являются откликом на определенные комбинации воздействия электромагнитных полей в диапазоне УФ-ИК. Реплики могут выстраиваться строго по горизонтали в обе стороны, двигаться по сложным траекториям, а также сохраняться какое-то время после прекращения воздействия полей на препарат ДНК. (Рис.1-4).

|
| |
Рис.1. Эффект волнового реплицирования ДНК и ближних окружающих предметов. Слева контроль, справа опыт. Второй вариант методов выявления волновых реплик ДНК.

|
|
Рис.2 (a). Дискретность и сложность траектории волновых реплик ДНК. (слева – исходное изображение, справа – контрастированное). Первый вариант методов выявления волновых реплик ДНК.

|

|

|
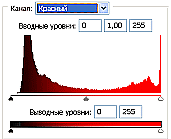
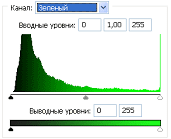
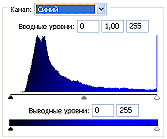
Рис.2 (б). Распределение яркостей изображения по каналам цветности: красный (R), зеленый (G), синий (B).

|

|



Рис 3 (б).Распределение яркостей по каналам цветности: красный (R), зеленый (G), синий (B).
|
|
|
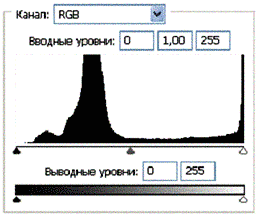
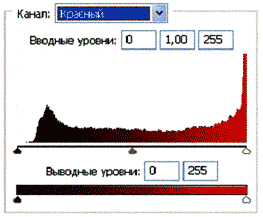
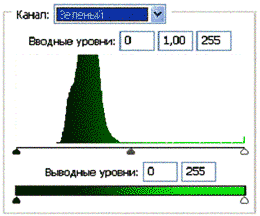
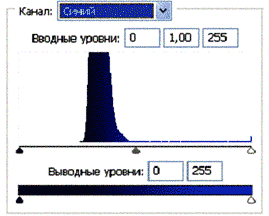
Рис 3.2. Гистограммы значений яркостей по каналам цветности: красный, зеленый, синий.
На контрольных снимках, когда источники воздействующих на биопрепарат физических полей не включены, данного явления не наблюдается. Также как нет эффектов при действующей аппаратуре, но без ДНК. За время экспозиции (1 сек.) в первом варианте экспериментов (см. Методы) волновые реплики ДНК перемещаются в пространстве и мультиплицируются, их траектория не предсказуема и дискретна. Генерация реплик происходит только при условии взаимной пространственной стационарности препарата ДНК, источников возбуждающих полей и фотоаппарата. Прикосновение к препарату ДНК (второй вариант экспериментов) вызывает изменение вектора распространения реплик в пространстве на диаметрально противоположное (слева направо), а затем исчезновение (Рис.5 (а, б, в)). Существенно, что в ряде случаев имеет место не только волновое мультиреплицирование препарата ДНК, но и окружающих предметов (Рис. 1, 5 (а, б)).
Для анализа цветного отпечатка на эмульсии фотопленки приведены изображения, полученные при сканировании пленки, как распределение яркостей в разных каналах цветности: красном (R), зеленом (G), синем (B): на Рис. 2(б), 3(б), 4(б). В изображении, представленном на Рис. 2 (а, б), видно, что почти вся яркостная шкала изображения реплик находится в красном канале цветности, в зеленом и синем они слабо различимы. Такое же превалирование яркостного диапазона красного канала хорошо видно на сопровождающих Рис.3 (б) гистограммах яркостей (Рис. 3 (в)) соответствующих каналов. Обращает на себя внимание именно та часть гистограммы, где расположено реплицированное изображение: участок яркостей в диапазоне значений от 128 до 255 – хорошо отличаемый пик значений в этой части гистограммы красного по сравнению с почти равномерным убыванием в такой же области гистограмм зеленого и синего каналов. Возможно, такое распределение яркостей может указывать на то, что сфотографированный дискретный трек есть опосредованная препаратом ДНК многократно повторенная реплика световой матрицы аппарата «Дюна-М» красного и инфракрасного цветов (позиция 8 на Рис. 8), на которой находится препарат ДНК в момент генерации реплик. Этот источник света представляет матрицу из 37 чередующихся диодов, из них красных – 21, инфракрасных – 16. В эксперименте препарат ДНК был помещен в пробирку эппендорф и перекрывал 5 диодов. Это коррелирует с 5-кратной продольной исчерченностью трека реплики (Рис. 3a).
Важным свойством волновых реплик ДНК является относительная долговременность их существования после выключения всех инициирующих физических полей (Рис. 4), но время жизни реплик может зависеть также от чувствительности фотопленки, фиксирующей реплики. А также от ее спектральной избирательности.

|
Рис. 4 (a). Волновая долгоживущая реплика ДНК |

|

|

|
Рис. 4 (б). Распределение яркостей по каналам цветности: красный, зеленый, синий.
|
|
|
|
Рис. 4 (в).Гистограммы. Распределение яркостей по каналам цветности: красный, зеленый, синий.
Первая слева в верхнем ряду: гистограмма RGB – общая (совмещенная).
Можно заметить существенное отличие в распределениях яркостей по каналам цветности у изображения волновой реплики ДНК и «фантома» этой реплики, остающегося после выключения источников света. Анализ изображений на Рис. 4 (б) и сопоставление его с гистограммой на Рис.4 (в) выявляет то, что изображение «фантома» реплики в красном канале становится размытым как по диапазону, так и по амплитудам значений яркостей. При этом в зеленом и синем каналах наблюдается сужение локализации значений в диапазоне от 70 до 120, виден четкий пик, что объясняет яркость реплики ДНК, ее оттенки и полутона, особенно, в зеленом канале.
|
Рис.5 (а). Момент механического воздействия на препарат ДНК. Второй вариант методов выявления волновых реплик ДНК. |

|
Рис. 5 (б). Сдвиг волновых реплик налево сразу после механического воздействия. Характерно появившееся резкое деление кадра по свето- и цветопередаче, не связанные со стабильной работой шторки фотоаппарата. |
|

|
Рис. 5 (в). Исчезновение эффекта образования реплик ДНК через 5-8 сек. после механического воздействия на препарат ДНК. При этом вся аппаратура, инициирующая реплики, остается включенной. |

Тип образующихся реплик в каждом конкретном случае в рамках используемых методов стохастический (1-й вариант методов) или детерминированный (2-й вариант) в пространстве-времени.
Одним из результатов по индуцированию волновых реплик окружающих предметов с помощью препарата ДНК по первому методу явилось мультиплицирование тройного изображения лампы БС (УФ-В) (Рис. 6).
Оригинал. |
|

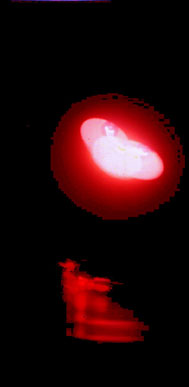
В качестве одного из контролей на способность генерировать волновые реплики использовали хлорид натрия (кристаллический); хлорид натрия (1М водный раствор); винная кислота в кристаллах; винная кислота рацемат (1М водный раствор); крахмал воздушно сухой; глицин кристаллический; кальциферол воздушно сухой; токоферол воздушно сухой; хлорофилл воздушно сухой; вода бидистиллированная; интерферон в смеси с Bacillus subtilis, воздушно сухие. Ни один из препаратов не дал реплик.
Методы

|
Рис. 7. Матрица с красными и инфракрасными диодами (аппарат «Дюна М» или «Дюна Т». http://argonet.ru/nar_lechebn_duna.htm) Содержит 37 диодов, из них красных – 21 ( |
Мы использовали две схемы экспериментов. Первая приведена на (Рис.9), вторая достаточно проста и видна напрямую на фото (Рис. 5(в)). Схемы также даны в18. Для получения и визуализации волновых репликативных отображений ДНК по первой схеме производят следующие операции. Посредством реле времени (позиция 3 на Рис.8) в разных сочетаниях включаются необходимые излучатели БС (УФ-В, представляющий лампу накаливания синего цвета «ЛОН БС 75», из стекла СЛ98-1) (позиция 5) и матрица с красными и инфракрасными диодами (Рис. 8)) (позиция 8 на схеме Рис. 8), а также лампа ртутная бактерицидная, или лампа Compact electronic CEST26E27 Black (УФ-С) (позиция 6), или БС (УФ-В) и МХТ-90 (холодный катод (позиция 4)). При включении источников света воздушно сухой препарат ДНК из селезенки быка (РЕАХИМ, марка А, НПО «Биолар») (около 100 мг в закрытой пластиковой конической пробирке «эппендорф» длиной 4см, диаметром в верхней части 0,9см) или (3мл водного раствора ДНК, 1мг/мл) находится в зоне действия излучателей (1мм-50см от источников света). Ход работы снимается на фотоплёнку Fuji 24-27 DIN. При этом работает осциллограф (позиция 2) с электродами, которые регистрируют электромагнитные поля в зоне проведения эксперимента и фиксируется усреднённый нормальный электромагнитный фон в помещении, определяемый по характеру синусоиды на осциллографе. Далее посредством реле времени через 10 мин. отключают излучатель УФ-С. Фотографически обнаруживается возникновение динамичных специфических волновых структур, невидимых глазами, но фиксируемых на фотопленке мультиплицированных реплик ДНК и ближних окружающих объектов, непосредственно связанных с фотонным воздействием на ДНК. Иначе говоря, происходит умножение количества отображений препарата ДНК, распределяемых в пространстве по сложным траекториям (1-й вариант методов) и по горизонтали (2-й вариант методов), а также отображений объектов, связанных с возбуждением ДНК.
Второй вариант методов получения и визуализации волновых репликативных отображений ДНК (Рис. 5(в)) состоит в следующем. Воздушно сухой препарат ДНК, 100мг, помещают в открытом виде в держатель из алюминиевой фольги. С интервалом 2-3 сек. включают лампу БС (УФ-В), лампу сompact electronic CEST26E27 Black (УФ-С) и аппарат «Дюна М». Через 5 мин. начинают фотосъемку на фотоплёнку Fuji 24-27 DIN. В этом варианте регистрируются реплики ДНК и ближних объектов, которые распространяются строго направо. При механическом воздействии на препарат ДНК вектор распространения реплик меняет своё направление на диаметрально противоположное, то есть налево. Затем через 5-8 сек. после механического воздействия, несмотря на то, что вся аппаратура, инициирующая реплики, остается включенной, реплики исчезают (или не фиксируются используемым типом фотопленки).
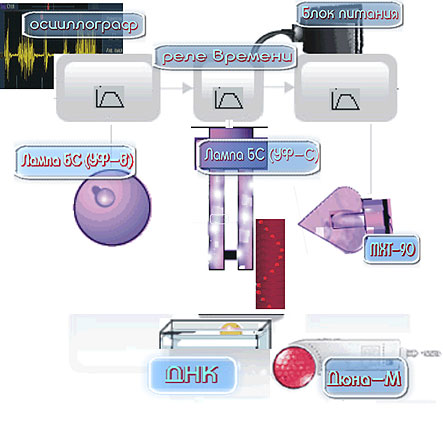
Рис. 8.
Обсуждение
Несмотря на успехи молекулярной генетики и клеточной биологии, до сих пор тонкие механизмы знаковых функций генетического аппарата остаются непонятными. Особенно ярко это проявилось после публикации работы, в которой команде Pruitt удалось изящно доказать, что законы Менделя в генетике не всегда выполняются, более того, поведение одного из генов не подчиняется здравому смыслу1. Это проявилось в том, что во взрослых растениях Arabidopsis фенотипически проявился нормальный ген Hothead, которого не было в исходных мутантных по этому гену семенах. Нормальный ген Hothead был у предков растения. Отсутствующий в хромосомах посеянных семян ген Hothead в 10% случаев заменил реальный мутантный ген hothead, когда эти семена дали взрослые растения. Объяснения этому феномену пока не дано. Высказано предположение, что нормальный ген хранился как его ревертазная РНК копия. Это слабое и уязвимое объяснение, не имеющее экспериментального доказательства. Необъяснимый с позиций классической генетики феномен возврата предкового гена поднял на поверхность целый ряд нерешенных кардинальных вопросов генетики и эмбриологии. Их можно кратко перечислить:
a/ воблирование 3-го нуклеотида в кодоне делает каноническую таблицу генетического кода ареной потенциальных ошибок в синтезе белков поскольку она автоматически даёт омонимии значащих дублетов в кодонах, когда пары одинаковых дублетов кодируют разные аминокислоты2; при этом третьим нуклеотидом кодона может быть любой из четырех, что постулировал F.Крик3; тем не менее, ошибки при выборе аминокислот чрезвычайно маловероятны, и это удивительный факт; b/ ситуация, когда 98% генома эукариот считают «мусором», в лучшем случае, помощником триплетного кода, или «кладбищем» вирусных геномов; c/ 2% кодирующей ДНК человека (около 40тыс. генов) оказалась весьма близкими таковым свиней, ослов, мух и даже E.coli; d/ гены по непонятным причинам и законам транспозицируются в 3-D континууме хромосом; e/ гены по непонятным причинам разбиты на интроны и экзоны; f/ матрично независимый синтез РНК последовательностей Q![]() репликазой бактериофага E. coli и такой же синтез РНК последовательностей РНК полимеразой бактериофага T7 E. coli4,5, которые нарушает канонический принцип чисто вещественной репликации ДНК
репликазой бактериофага E. coli и такой же синтез РНК последовательностей РНК полимеразой бактериофага T7 E. coli4,5, которые нарушает канонический принцип чисто вещественной репликации ДНК![]() РНК; g/ фантомный листовой эффект, когда часть живого листа растения воспроизводит его целый образ в режиме газо-разрядной (Кирлиановской) визуализации6,7; h/ непонятные по механизмам дистантные функции селекторных генов морфогенеза, когда их продукты синтезируются в одном месте, а действие сразу проявляется в другом, далеко отстоящем месте развивающегося эмбриона.
РНК; g/ фантомный листовой эффект, когда часть живого листа растения воспроизводит его целый образ в режиме газо-разрядной (Кирлиановской) визуализации6,7; h/ непонятные по механизмам дистантные функции селекторных генов морфогенеза, когда их продукты синтезируются в одном месте, а действие сразу проявляется в другом, далеко отстоящем месте развивающегося эмбриона.
Все эти и другие мало понимаемые проявления знаковых функций хромосом заставляют думать и доказывать, что генетическая память имеет иные атрибуты, существенно дополняющие белковый код. Вероятно, эти атрибуты имеют волновую природу. Именно эти противоречия и неполнота модели триплетного белкового кода для объяснения кодирования пространственно-временной структуры многоклеточных биосистем заставила нас обратить внимание на работу генетического аппарата как системы высоко организованных знаковых излучений электромагнитных и акустических полей8,9,10,11. Вероятно, одним из проявлений волновых знаковых функций ДНК являются обсуждаемые здесь результаты нашего исследования по волновым репликам ДНК. Первые свидетельства, что ДНК способна к волновому типу памяти были получены нами в 1985 г. Мы обнаружили феномен так называемой ДНК фантомной памяти (DNA phantom effect)12и подробно описали его в работе8. Возможно, он имеет прямое отношение к излагаемым здесь результатам. ДНК фантомный эффект проявляется следующим образом. При работе с ДНК, как линейным полимером, с использованием метода динамического лазерного светорассеяния (ДЛС), в кюветном отделении спектрометра остаются некие гипотетические структуры (объекты), специфическим образом рассеивающие свет после удаления кюветы с препаратом ДНК. Именно это явление было названо ‘DNA phantom effect’. Контрольные замеры, до помещения ДНК в спектрометр, дают только фоновое светорассеяние. Продувание кюветного отделения прибора газообразным азотом приводит к исчезновению фантома, однако он вновь регистрируется спустя 5-7 минут. Фактически близкий эффект наблюдали Allison et. al.13, которые также использовали ДЛС для изучения рестриктных фрагментов ДНК. Они обнаружили аномальное светорассеяние таких фрагментов. Авторы определили этот феномен как ‘mimicking the effect of dust’, то ест как эффект, имитирующий пыль. В их исследовании этот эффект не являлся результатом пылевых загрязнений, но следствием влияния ‘объекта’, который вел себя как пылеподобные ‘частицы’, рассеивающие свет. Поведение таких ‘частиц’ задает такие характеристики светорассеяния, которые не согласуются с классическими моделями процесса светорассеяния для линейных полимеров14,15,16. Между экспериментами8,12,13есть общие моменты, связанные с тем, что на препараты ДНК воздействовали светом в видимой области – 632,8нм и 488нм. То же самое имеет место при инициации волновых реплик ДНК в данной работе. В образовании волновых реплик ДНК доминирующую функцию берёт на себя, вероятно, тот излучатель, длина волны которого совпадает с длиной волны поглощения ДНК. Это источник УФ света.Мы не можем исключить, что в экспериментах8,12,13возникали волновые реплики ДНК, которые искажали классическое светорассеяние для линейных полимеров. Мы не можем также исключить, что волновые реплики ДНК могут играть определенную роль в процессах волновых генетических коммуникаций между клетками в онтогенезе и во взрослом состоянии организмов.
Для волновых реплик ДНК in vitro свойственен квази генетический процесс: ДНК волновым путем копирует самою себя и объекты (устройства, оборудование), вовлеченные в инициацию реплицирования (Рис. 1-3, 5 (a), (b); 6 (a), (b)). Этот момент поведения генетического аппарата in vivo у многоклеточных организмов является одним из ключевых в нашей модели волнового генома8,9,10,11. В соответствии с этой моделью, каждая клетка и вся биосистема в целом осуществляет непрерывное сканирования своего генетико-метаболического состояния. Иными словами, ДНК за счет собственных когерентных излучений in vivo поляризационно-голографическим образом19считывает самою себя и внутриклеточное метаболическое пространство в диапазоне 250-800нм, то есть копирует, создает волновые реплики своего структурно-функционального статуса в каждый момент времени. Возможно, диапазон частот гораздо шире, но технически пока возможна регистрация только указанного спектра частот. Такие реплики образуют так называемое спутанное (entanglement) состояние и нелокально (и мгновенно) информационно объединяют биосистему20.
Вместе с тем, мы должны подчеркнуть, что обнаруженные эффекты образования волновых реплик ДНК нуждаются в более точной экспериментальной постановке. Очень многое в этой части остается неясным и поэтому исследования продолжаются.
ТЕОРЕТИЧЕСКОЕ ПРИЛОЖЕНИЕ
В течение длительного времени физика оперировала четырьмя взаимодействиями, — сильными и слабыми взаимодействиями, электромагнитными и гравитационными полями. С помощью этих взаимодействий удавалось объяснить почти все известные явления природы и многочисленные лабораторные эксперименты. Однако постепенно стала накапливаться экспериментальная феноменология, которую не удавалось объяснить с помощью указанных взаимодействий21. Методологически было понятно, что такое положение свидетельствовало о неполноте наших знаний о природе. Для понимания экспериментальной феноменологии надо было сформулировать новую научную парадигму. Первые шаги в этом направлении были сделаны Э.Картаном, который постулировал существование в природе полей, порождаемых плотностью углового момента вращения22. Эти поля были названы им полями кручения или торсионными полями. Многочисленные исследования в этом направлении в 70-е годы ХХ столетия привели к созданию Теории Эйнштейна – Картана (ТЭК), которая связывала природу кручения с гравитацией. Позже было показано, что торсионные поля порождаются спином, в отличие от электромагнитных полей, порождаемых зарядом, и гравитационных полей, порождаемых массой. В этом случае торсионные поля рассматриваются как самостоятельная физическая реальность, как пятое взаимодействие, «пятая сила».
В 80-е годы ХХ столетия А.Е.Акимовым была построена фитонная модель физического вакуума21, которая в период до создания теории физического вакуума Г.И.Шиповым23позволила понять физические механизмы, связанные с действием торсионных полей. При построении моделей физического вакуума было признано целесообразным вернуться к электронно-позитронной модели П.Дирака в несколько измененной ее интерпретации. Учитывая, что вакуум определяется как состояние без частиц, и, исходя из модели классического спина Я.Френкеля и И.Тернова24,25как кольцевого волнового пакета (следуя терминологии Белинфанте — циркулирующего потока энергии), будем рассматривать вакуум как систему из кольцевых волновых пакетов, соответствующих электронам и позитронам, а не собственно электронно-позитронным парам, несущим информацию о свойствах этих частиц (рис. 1).
При сделанных предположениях нетрудно видеть, что условию истинной электронейтральности такого полевого электронно-позитронного вакуума будет отвечать состояние, когда кольцевые волновые пакеты электрона и позитрона будут вложены друг в друга. Если при этом спины этих вложенных кольцевых пакетов противоположны, то такая система будет самоскомпенсирована не только по зарядам, но и по классическому спину и магнитному моменту. Так как масса покоя волновых пакетов равна нулю, то такие системы будут обладать гравитационной нейтральностью. Такую систему из вложенныхкольцевых волновых пакетов будем называть фитоном. Фитонный континуум образует материальную среду – физический вакуум. Рассмотрим наиболее важные в практическом отношении случаи возмущения разными внешними источниками физического вакуума в рамках построенной модели.
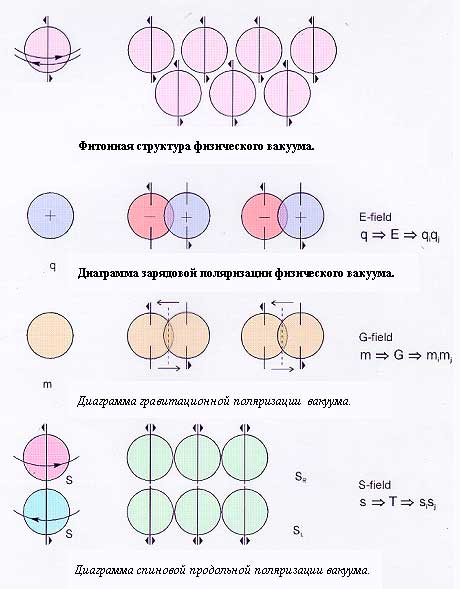
Рис.1. Фитонная структура физического вакуума и его поляризационные состояния.
1. Пусть источником возмущения является заряд — q. Если вакуум имеет фитонную структуру, то действие заряда будет выражено в зарядовой поляризации физического вакуума. Это состояние фиксируется как электростатическое или электромагнитное поле. Этот случай хорошо известен в квантовой электродинамике. В частности, Лэмбовский сдвиг традиционно объясняется через зарядовую поляризацию электронно-позитронного физического вакуума.
2. Пусть источником возмущения является масса — m. В отличие от предыдущего случая, когда мы столкнулись с общеизвестной ситуацией, здесь будет высказано гипотетическое предположение. Возмущение физического вакуума массой m будет выражаться в симметричных колебаниях элементов фитонов вдоль оси на центр объекта возмущения. Такое состояние физического вакуума может быть охарактеризовано, как гравитационное поле (G-поле).
3. Пусть источником возмущения является классический спин — s. Будем предполагать, что действие классического спина на физический вакуум будет заключаться в следующем. Если источник имеет спин, то спины фитонов, которые совпадают с ориентацией спина источника, сохраняют свою ориентацию. При этом спины фитонов, которые противоположны спину источника, под действием источника испытают инверсию. В результате физически вакуум перейдет в состояние поперечной спиновой поляризации. Это поляризационное состояние можно интерпретировать как спиновое поле (S — поле) — поле порождаемое классическим спином, т.е. торсионное поле, — в другой интерпретации дальнодействующее спинорное поле, если учесть, что торсионное поле может быть представлено через пару спинорных полей. Сформулированный подход созвучен представлениям о полях кручения, как конденсате пар фермионов.
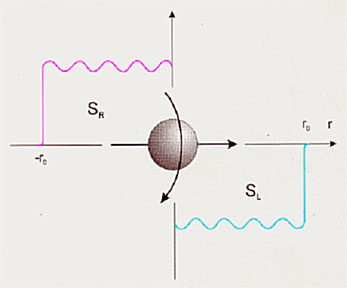
Поляризационные спиновые состояния SR и SL противоречат запрету Паули. Однако согласно концепции М.А.Маркова26, при плотностях порядка планковских, фундаментальные физические законы могут иметь другой, отличный от известных вид.
В соответствии с изложенным подходом можно говорить, что единая среда – физический вакуум может находиться в разных фазовых, точнее, поляризационных состояниях — EGS состояниях. Эта среда в состоянии зарядовой поляризации проявляет себя как электромагнитное поле (Е). Эта же среда в состоянии спиновой продольной поляризации проявляет себя как гравитационное поле (G). Наконец, та же среда — физический вакуум в состоянии спиновой поперечной поляризации проявляет себя как спиновое (торсионное) поле (S). Таким образом, ЕGS — поляризационным состояниям физического вакуума соответствуют EGS -поля.
В начале 80-х годов в работах А.Е.Акимова было предсказано и экспериментально подтверждено существование особого класса торсионных полей — электроторсионных полей21. В середине 90-х годов автор теории физического вакуума Г.И.Шипов дал строгое на фундаментальном уровне обоснование электроторсионных полей22.
Модель поляризационных состояний физического вакуума позволила установить весьма важное обстоятельство. Уже отмечалось, что торсионное поле фиксируется в случаях, когда физический вакуум переходит в состояние спиновой поперечной поляризации. Однако можно сформулировать более общий подход, опираясь на критерии синергетики. Будет считать, что торсионное поле фиксируется всегда, когда физический вакуум находится в спиново неравновесном состоянии.
Нетрудно видеть, что при зарядовой поляризации физического вакуума зарядовое расщепление фитонов приводит и к спиновому пространственному расщеплению. В результате спины оказываются некомпенсированными, что будет равносильно появлению торсионной компоненты в электромагнитном поле. Если гравитационные и торсионные поля проявляются в «чистом виде», то электромагнитные поля всегда содержат торсионную компоненту, что является важным фундаментальным фактом. Торсионное поле наблюдается как в электростатическом поле, так и в электромагнитном излучении.
С изложенных позиций нетрудно предположить, что при электромагнитном или звуковом воздействии на субстрат ДНК создаётся его зарядовая неравновесность. Следствием этого является возникновение электроторсионного поля. В рассмотренных выше экспериментах неотвратимо практически все объекты становились такими источниками. При этом первичным объектом возмущения был субстрат ДНК, который своим электроторсионным полем индуцировал возмущение других объектов эксперимента.
| a) |
|
| б) |
|
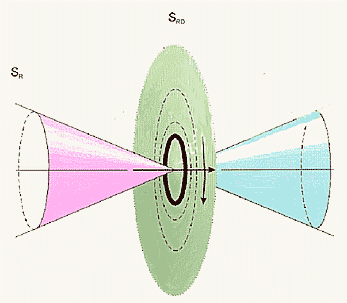
Рис.2. Диаграмма направленности торсионного поля спинирующего источника (а)
и зависимость интенсивности статического торсионного поля от расстояния (б).
Теперь рассмотрим диаграмму направленности источника торсионного поля, в частном случае, — объект со спином или с угловым моментом вращения для макроскопических тел (рис. 2). На расстояниях больших размера источника электроторсионного поля это поле имеет периодическую структуру. Поэтому на каждом интервале такой периодической структуры будет отображение источника торсионного поля. В зависимости от знака поля внешнего воздействия будет отображаться, либо правая, либо левая часть таких периодических структур. Возможно, именно этот эффект мы наблюдаем на Рис. 5 (а, б) (экспериментальная часть), где эффект правых реплик ДНК и ее окружения меняется на левые реплики при механическом воздействии на препарат ДНК.
При этом остаётся вопрос, как это всё может фиксироваться на фотографиях, если учесть, что фотоплёнка фиксирует изображения фотохимическим способом, т.е. за счёт электромагнитных воздействий, а сами исходные структуры имеют торсионную природу. Возможность фиксации на плёнках торсионных пространственных структур связана со следующими факторами. В фотографических устройствах (фотоаппаратах с фотоплёнкой) фотоны, попадая в эмульсию, вызывают фотохимические реакции. В результате при последующих химических реакциях проявления и закрепления на фотоплёнке появляется изображение. При действии торсионных излучений на фотоактивные вещества эмульсии фотоплёнки изменяется спиновое состояние этих веществ. Результат изменения спинового состояния атомов приводит к химическим изменениям как следствие изменения взаимодействия валентных электронов, которое подобно тем изменениям, которые наблюдаются при действии на эти вещества фотонов. Это и позволяет получать фотографии пространственных структур, которые обсуждаются в данной статье.
Физическая интерпретация наблюдаемых явлений, представленных выше, базировалась на важных, но частных эффектах проявления торсионных полей. Рассмотрим указанную экспериментальную феноменологию с позиций основных свойств торсионных полей. Укажем на наиболее важные свойства торсионных полей.
1. Источником торсионных полей является классический спин или макроскопическое вращение. Торсионные поля могут порождаться кручением пространства или порождаться как следствие возмущения физического вакуума, которое имеет геометрическую или топологическую природу, а также возникать как неотъемлемая компонента электромагнитного поля. Торсионные поля могут самогенерироваться.
Во всех указанных случаях речь идет о торсионных полях, порождаемых на уровне вещества. Однако согласно теории физического вакуума существуют первичные торсионные поля, которые порождаются Абсолютным Ничто. Подобно тому, как исходный материал мира вещества — элементарные частицы — рождаются из физического вакуума, в свою очередь физический вакуум рождается из первичного торсионного поля.
2. Квантами торсионного поля являются тордионы. Есть основания считать, что тордионами являются низкоэнергетические нейтрино с энергией порядка единиц эВ.
3. Так как торсионные поля порождаются классическим спином, то и при их воздействии на те или иные объекты у этих объектов в результате воздействия может измениться только их спиновое состояние (состояние ядерных или атомных спинов, а также спинов валентных электронов).
4. Торсионные поля обладают аксиальной симметрией относительно своего источника.
5. Торсионные поля (Т), порождаемые классическим спином, могут быть аксиальные (Тa) и радиальные (Тr). Каждое из этих торсионных полей может быть правое (ТaR, ТaL) или левое (TaL, ТrL). В общем случае торсионные поля обладают аксиальной симметрией.
6. Одноименные торсионные заряды (одноименные классические спины (SRSR или SLSL) притягиваются, а разноименные (SRSL) — отталкиваются.
7. Стационарный спинирующий объект создает статическое торсионное поле. Если у спинирующего объекта есть какая-либо неравновесность: изменение угловой частоты, наличие у объектов прецессии, нутации или моментов более высокого порядка, неравномерного распределения масс относительно оси вращения для массивных объектов, то такой динамический спинирующий объект создает волновое торсионное излучение.
8. Статическое торсионное поле имеет конечный радиус действия rо, на интервале которого интенсивность торсионного поля слабо варьирует. Условно по аналогии с электромагнетизмом, хотя физика процессов здесь другая, этот интервал rо можно назвать ближней зоной. Волновое торсионное излучение не ограничено интервалом rо и его интенсивность не зависит от расстояния.
9. Средой, через которую распространяются торсионные излучения, является физический вакуум. По отношению к торсионным волнам физический вакуум ведет себя как голографическая среда. В этой среде торсионные волны распространяются через фазовый портрет этой голограммы. Этот основополагающий физический фактор объясняет информационный (не энергетический) характер передачи сигналов, а также сверхсветовую скорость передачи сигналов.
10. Для торсионных полей потенциал тождественно равен нулю, что соответствует их неэнергетическому характеру. Это второй фактор, который определяет почему торсионные сигналы (воздействия) передаются информационно, а не энергетически, т.е. без переноса энергии.
11.Константа спин-торсионных взаимодействий для статических торсионных полей с кручением Картана по существующим оценкам меньше, чем 10-50, т.е. для таких полей невозможно существование наблюдаемых эффектов. Для волновых торсионных полей с кручением Картана (динамическое кручение) константа спин-торсионных взаимодействий теоретически не ограничена. Для торсионных полей с кручением Риччи или Вайценбека нет органический на величину константы взаимодействий, а, следовательно, и на интенсивность проявления этих полей. Для торсионных полей с кручением, порождаемых как компонента электромагнитных полей (электроторсионные взаимодействия) константа взаимодействий имеет порядок 10-3 –10-4.
12. Так как константа электроторсионных взаимодействий (10-3-10-4) чуть меньше константы электромагнитных взаимодействий (1/137~10-2), то в естественных условиях такие торсионные воздействия могут вызвать наблюдаемые изменения или фиксироваться как наблюдаемые сигналы только в тех объектах, в которых есть неравновесные состояния, ослабляющие электромагнитные связи.
13. Торсионные поля проходят через природные среды без потерь. Это является естественным фактором, если учесть, что квантами торсионных полей являются нейтрино. Полезно отметить, что без связи с торсионными полями советскими физиками более десяти лет назад было показано, что спиновые возмущения в спиновой среде распространяются так, что их нельзя экранировать27.
14. Скорость торсионных волн теоретически равна бесконечности. Сверхсветовые скорости не являются чем-то необычным для физики. Они присутствовали в теории гравитации Ньютона, они составляют основы концепции тахионов. Сверхсветовые скорости впервые наблюдались экспериментально Н.А.Козыревым28, позже подтвержденные двумя другими коллективами29,30, а на квантовом уровне Цойлингером31.
15. Все тела живой и неживой природы состоят из атомов, большинство которых обладают ненулевыми атомными или ядерными классическими спинами. Учитывая, что все тела находятся в магнитном поле Земли, за счёт магнитных моментов ядер и атомов, которые являются следствием наличия указанных классических спинов, возникает прецессия, которая порождает волновое торсионное излучение. Таким образом, все тела обладают собственными торсионными полями (излучениями).
16. Так как разные тела обладают разным набором химических элементов, разным набором химических соединений с разной стереохимией, и разным пространственным распределением в телах этих атомов и химических соединений, то все тела обладают строго индивидуальными, характеристическими торсионными полями (излучениями).
Из перечисленных свойств торсионных полей становится понятно, почему в экспериментах с ДНК фиксируются пространственные структуры торсионных полей характеристической формы для разных объектов. Становится ясно, за счёт чего в экспериментах с ДНК наблюдаются голографические признаки, в частности, нелокальные взаимодействия, а сами даже дальние взаимодействия реализуются мгновенно.
Литература
- Lolle S.J., Victor J.L., Young J.M., Pruitt R.E. Genome-wide non-mendelian inheritance of extra genomic information in Arabidopsis. Nature, 434, 505-509 (2005).
- Lagerkvist U., «Two out of Three»: an alternative method for codon reading. Proc. Natl.Acad.Sci.USA, 75, 1759- 1762 (1978).
- Francis Crick. A personal View of Scientific Discovery. Basic Books, Inc., Publishers, New York. Translate into Russian: Institute of computer researches, Moscow-Ijevsk (2004), p. 98.
- Biebricher C.K., Eigen M., Luce R. Product analysis of RNA Generated de novo by Qb Replicase. J.Mol.Biol., 148, 369-390 (1981).
- Biebricher C.K., Luce R., Template-free generation of RNA species that replicate with bacteriophage T7 RNA polymerase. The EMBO Journal, 15, Issue 13, 3458-3465 (1996).
- Choudhury G.K., Kejarival P.C., Chattopadhyay A. The devices for generation of phantom leaf effect. J.Inst.Eng. (India) 60, Pt.EL3, 61-66; 67-73 (1979).
- Gariaev P.P., Junin A.M. Phantom leaf effect. Myth or reality? Energy 10, 46-52 (1989). In Russian.
- Gariaev P.P. Wave based genome. Monograph. Moscow. Ed. Obshestv. Pol’za. 279p. (1994). In Russian.
- Gariaev P.P. Wave genetic code. Monograph. Moscow. Ed. Izdatcentr. 108p. (1997). In Russian.
- Prangishvili I.V., Gariaev P.P., G.G.Tertishny, V.V.Maximenko, A.V.Mologin, E.A.Leonova, E.R.Muldashev. Spectroscopy of radiowave radiation of localized photons: path to quantum nonlocality of bioinformation processes. Sensors and systems, 9 (18), 2-13 (2000). Russian Academy of sciences. In Russian.
- Gariaev P.P., Birshtein B.I., Iarochenko A.M, Marcer P.J., George G. Tertishny G.G., Leonova E.A., Kaempf U., The DNA-wave biocomputer. «CASYS» – International Journal of Computing Anticipatory Systems (ed. D.M.Dubois), Liege, Belgium, 10, 290-310, (2001).
- Gariaev P.P., Chudin V.I., Komissarov G.G., Berezin A.A., Vasiliev A.A., Hologrphic Associative Memory of Biological Systems, Proceedings SPIE — The International Society for Optical Engineering. Optical Memory and Neural Networks, 1621, 280- 291. USA. (1991).
- Allison S.A., Sorlie S.S., Pecora R., Brownian Dynamics Simulations of Wormlike Chains, Dynamic Light Scattering from a 2311 Base Pair DNA Fragment. Macromolecules, 23, 1110-1118, (1990).
- Hagerman, P. J., and B. H. Zimm., Monte Carlo approach to the analysis of the rotational diffusion of wormlike chains. Biopolymers, 20,1481–1502, (1981).
- Zimm B H., Dynamics of polymer molecules in dilute solution: viscoelasticity, flow birefringence and dielectric loss, J. Chem. Phys., 24, 269-78, (1956). General Electric Research Laboratory, Schenectady, NY.
- Rouse P E., A theory of the linear viscoelastic properties of dilute solutions of coiling polymers, J.Chem.Phys., 21, 1272-80, (1953). Tertyshnii G.G., Gariaev P.P., Aksenov V.A., Leonova E.A., Fomchenkov S.V., The formalism of endogenous polarization/holographic managing processes in organisms. Consciousness and a physical reality, 9, number 4, 44-50 (2004). In Russian.
- Prangishvili I.V., Gariaev P.P., Tertishnii G.G., Maksimenko V.V., Mologin A.V., Leonova E.A., Muldashev E.R., Spectroscopy of radiowave radiations of the localized photons: an output on quanum nonlocal bioinformation processes. Sensors and Systems, 9 (18), 2-13 (2000). In Russian.
- Заявление о выдаче патента в Федеральный институт промышленной собственности России от 20.10.2005. Регистрационный номер 2005132403. Входящий номер 036308.
- Tertyshnii G.G., Gariaev P.P., Aksenov V.A., Leonova E.A., Fomchenkov S.V., The formalism of endogenous polarization/holographic managing processes in organisms. Consciousness and a physical reality, 9, number 4, 44-50 (2004). In Russian.
- Prangishvili I.V., Gariaev P.P., Tertishnii G.G., Maksimenko V.V., Mologin A.V., Leonova E.A., Muldashev E.R., Spectroscopy of radio wave radiations of the localized photons: an output on quanum nonlocal bioinformation processes. Sensors and Systems, 9 (18), 2-13 (2000). In Russian.
- А.Е.Акимов. Эвристическое обсуждение проблемы поиска новых дальнодействий. EGS-концепции. МНТЦ ВЕНТ, препринт №7А, Москва, 1992.
- Е.Cartan On the Generalization of the Notion of the Curvature of Riemann and spaces whit Torsion» (Comptes Rendus Acad. Sci, vol 174, 1922, p.593-595),
- Г.И.Шипов. Теория физического вакуума. М., Наука, 1997, с.450.
- М.М.Тернов, В.А.Бордовицын. О современной интерпретации классической теории спина Я.М.Френкеля. УФН, 1980, т. 132, вып.2, с.345.
- Б.Г.Багров, Б.А.Бордовицын. Классическая теория спина. Известия ВУЗ, сер. Физика, 1980,Ш, с.67.
- Markov M.A. Very Early Universe. Proc. Of the Nuffield Workshop. Eds. Gibbson G.W., Hawking S.W., Siklov S.T. — Cambridge, 1988,p.353.
- Л.Б.Окунь. Физика элементарных частиц. Наука, ГР ФМЛ, М., 1988, с.272.
- Козырев Н.А., Насонов В.В. О некоторых свойствах времени, обнаруженных астрономическими наблюдениями. — Проблема исследования Вселенной, 1980, вып.9, с.76.
- А.Е.Акимов, А.Ф.Пугач. К вопросу о возможности обнаружения торсиопных воли астрономическими методами. МНТЦ ВЕНТ, М., 1992, препринт №25, с.19.
- М.М.Лаврентьев, И.А.Еганова, М.К.Луцет, С.Ф.Фоминых. О дистанционном воздействии звезд на резистор. Доклады АН СССР, 1990, т.314, вып.2, с.352.
- Dik Bouwmeester, Jian-Wei Pan, Klaus Mattle, Manfred Eibl,
Harald Weinfurter & Anton Zeilinger. Experimental quantum teleportation, Nature, v.390, N11, pp.575-579, December 1997.
П.П.Гаряев, Валтрауд Вагнер, Е.А.Леонова-Гаряева, А.Е.Акимов, И.А.Сухоплюева, Э.Р.Мулдашев Волновые репликативные отображения ДНК и ее ближайшего окружения // «Академия Тринитаризма», М.,
|
|