|
|
|
Назарова А.Ф.
Мы обнаружили европеоидные частоты генов белков и ферментов крови (1) в семи популяциях алтайцев. Вычисление генетических расстояний 11 человеческих популяций Европы, Азии и Америки, а именно саамов (лопарей), ненцев, нганасан, эвенков, якутов, монголов, алтайцев, русских, финнов, немцев и американских индейцев по частотам 28 аллелей белков, ферментов и групп крови показало определенную близость по наследственным признакам европеоидов и северных монголоидов (Табл.1). По данным матрицы генетических расстояний перечисленных выше человеческих популяций Европы, Азии и Америки мы построили дендрограмму родства популяций (Рис. 1), согласно которой мы полагаем, что отделение от общей праазиатской популяции предков американских индейцев произошло ранее всего (2).
Предположительно это происходило около 50 тыс. лет назад – это время разделения европеоидов и монголоидов по данным Неи (3), вычислившего генетические расстояния крупнейших человеческих рас по большому числу генетических маркеров. Следующими, согласно дендрограмме (Рис.1) от предковой популяции, обитавшей в регионе Южной Сибири и (или) в соседних регионах Центра Азии, отделились предки саамов. Оставшиеся популяции дендрограммы далее разделяются на два кластера, в один из которых входят два субкластера: ненцев, нганасан и эвенков, а в другой – якутов, монголов и алтайцев; и во второй кластер входят популяции современных европеоидов – русских, немцев и финнов (2). Такое родство европеоидов и северных монголоидов подтверждается данными о полиморфизме митохондриальной (mt)ДНК, которые обнаружили наличие одинаковых гаплогрупп у алтайцев и европеоидов Европы (4) – гаплогруппы H,J,K,T,U,V,W,F, у саамов эвенков и американских индейцев (5)–гаплогруппа V, и у алтайцев и американских индейцев (6)- гаплогруппы A, B, C,D. Наконец, обнаружение идентичной гаплогруппы X у европеоидов и у американских индейцев(7) подтвердило определенную общность происхождения этих двух групп человечества, причем методы молекулярной биологии показали, что эта гаплогруппа X у индейцев не привнесена контактами с европейцами уже после открытия Америки Колумбом, а является древней.
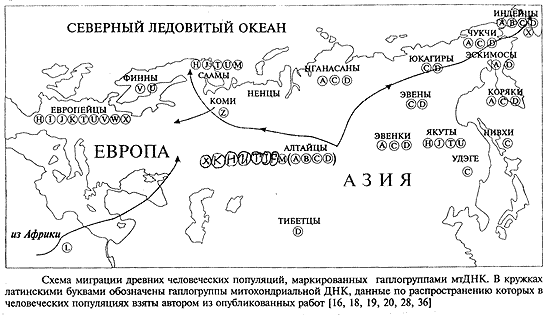
На рис.2 мы изобразили возможный путь миграции, начиная со среднего палеолита, человеческих популяций, маркированных гаплогруппами митохондриальной ДНК(4-7),от места их дифферен-циации в центре Азии до мест их теперешнего обитания.
Миграция древних человеческих популяций могла быть вызвана изменениями климата и обусловленных этим миграциями животных, на которых охотились люди палеолита. Предки американского северного оленя (карибу) и предки бизонов в палеолите обитали в регионе Южной Сибири (8), и с изменением климата продвинулись на крайний Северо-Восток Сибири, а потом перешли по существовавшему тогда Берингийскому перешейку в Америку.За ними шли древние племена америндов. Об общей тенденции представителей животного царства к миграциям в древности с Юга Сибири на ее Север говорят данные о миграциях беспозвоночных (кольчатых червей), и о миграциях птиц, приведенные в книге (9).
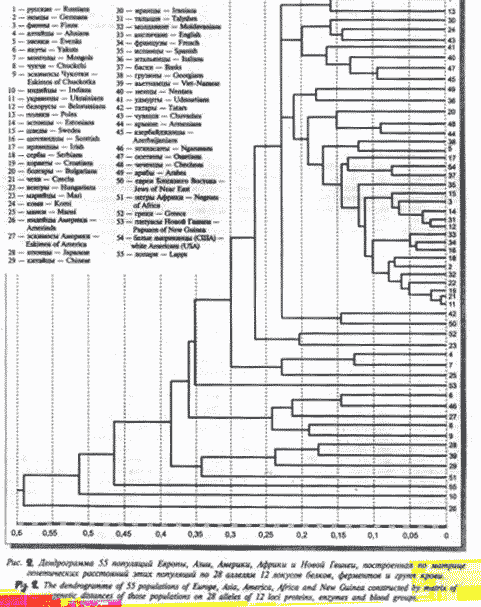
Мы вычислили матрицу генетических расстояний 55 человеческих популяций, принадлежащих к четырем большим человеческим расам, и обитающим в Европе, Азии, Америке, Африке и Океании (10). По данным этой матрицы мы построили эволюционную дендрограмму этих 55 популяций (Рис. 3). Из этой дендрограммы видно, что первыми отделяются, как и в дендрограмме на Рис.1, американские индейцы, затем саамы. К русским ближе всего оказались из славянских народов поляки, как и при вычислении нами генетических расстояний по меньшему числу генетических локусов (11).В одном большом кластере с русскими оказались иранцы, коми, чуваши, удмурты, ненцы,и субкластер осетин и азербайджанцев. Есть мнение, что русские происходят от древнеиранских скифских племен, мигрировавших ранее по Азии. Геродот именовал племена, обитавшие в его время на территории Восточной Европы, «скифами-пахарями». Племена же финно-язычных народов меря и мурома, близких к коми, чувашам и удмуртам, и обитавшие еще в I-м тысячелетии н.э. на месте Мос-ковской, Владимирской и Ярославской областей, полностью вошли в состав русского этноса и растворились в нем (Т.И.Алексеева, 1973) (13). Итак, близость русских к финским и иранским популяциям понятна.
Следующий большой кластер (Рис.3) начинается субкластером арабов и итальянцев, далее идет субкластер болгар, чеченцев и армян- это потомки племен, заселявших регион Кавказа и Закавказья; близость армян и чеченцев подтверждается близостью армянского и вайнахских языков (С.А.Старостин) (14). Следующий субкластер- ветви грузин и эвенков, их близость связана с тем, что предки грузин также кочевали в древности по Северной Азии; европеоидные частоты генов у эвенков мы обнаружили еще в 1984 г.(12). Вхождение басков и белых североамериканцев в один субкластер объясняется их общим кельтским субстратом. В соседние недавно разделившиеся субкластеры входят шведы, финны, эстонцы, талыши и белорусы. Талыши- это ираноязычный этнос Закавказья, изученный нами впервые в генетическом аспекте (15), его близость к северным европеоидам может указывать на пути миграции из Азии в Европу. Субкластер англичан, французов и шотландцев объединяется общим кельтским субстратом. В один субкластер входят немцы и сербы, и в три соседние субкластера- остальные народы Центральной и Юго-Восточной Европы: молдаване, венгры, хорваты, чехи и украинцы.
Последний субкластер этого большого кластера, начинавшегося ветвью русских,- греки и марийцы; греки – потомки древних европеоидов- ахейцев, в древности также, вероятно, мигрировали из Азии в Европу, как и марийцы — один из финских народов, чья праро-дина была в Азии (10). Далее идут азиатские субкластеры, первый их которых- алтайцы и монголы, а также манси- народ угорской группы. Вторая ветвь огромного кластера, включающего все европеоидные народы и субкластер переходных популяций –алтайцев, монголов и манси — это папуасы Новой Гвинеи. Следующим идет субкластер якутов и нганасан, а также эскимосов Америки, потом субкластер чукчей и эскимосом Чукотки, потом субкластер японцев и вьетнамцев, а также китайцев.Вторая ветвь кластера монголоидов Восточной Азии – это негры Африки. Итак, внизу дендрограммы остается ветвь лопарей (саамов), и ветвь индийцев, которые имеют сложное происхождение, являясь результатом метисации веддоидных племен и племен древних европеоидов.
Самая нижняя ветвь дендрограммы – это американские индейцы, предки которых первыми отделились от общей праазиатской популяции в среднем палеолите.
Есть антропологические данные, говорящие о нахожденнии европеоидов в Азии в палеолите. Так, академик В.П.Алексеев(16) писал об обнаруженных в пещере Дуньдянь в Китае черепах времен палеолита с европеоидными признаками. Писал он и о европео-идных признаках людей палеолита стоянки Сунгирь в Восточной Европе, появившихся там в более позднее время (25 000 лет назад).
Российский антрополог Т.В Томашевич обнаружила градиент распределения частот надглазничных каналов черепа человека (Табл.2), (17). В надглазничных каналах проходят надглазничные и надблоковые артерий и вен — ветвей глазничной артерии. В исключительных случаях в них проходят надглазничная и надблоковая ветви лобного нерва. Как видно из данных Табл. 2, у саамов, американских индейцев, а также северных монголоидов- якутов, юкагиров, эвенков, чукчей, эскимосов наиболее высокая частота встречаемости надглазничных каналов, что возможно, вызвано их палеоазиатским происхождением. У кетов, юкагиров, русских, манси, осетинов, армян частота надглазничных каналов очень близка и варьирует в пределах 30-38 %, это может быть подтверждением их общего азиатского происхождения. У монголоидов Восточной Азии – японцев, корейцев, китайцев, а также у монголов частота надглазничных каналов 43-48%. Интересно, что у древних египтян, этрусков, древних славян (полян киевских) и индийцев Уттар Прадеш частота каналов практически одинакова и составляет 25-28%, чуть выше она у словен новгород-ских; это также может быть связано, с происхождением этих народов из древней палеоазиатской популяции.
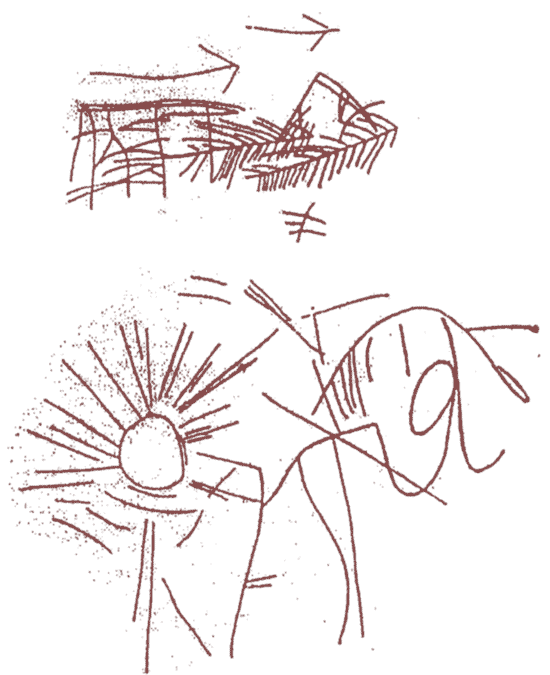
Отечественный шумеролог А.Г.Кифишин расшифровал петроглифы, обнаруженные археологами В.Е.Ларичевым и А.П.Окладниковым в Прибайкалье.т Как известно, академик А.П.Окладников опубликовал петроглифы, обнаруженные на берегах Лены, Байкала,Амура и на Алтае. Ларичев и Окладников исследовали археологическую культуру Мальты в Иркутской области, датируемой 20 000 лет до н.э. По мнению Кифишина, обнаруженная археологами близ села Суон-Тит на р. Алдан наскальная надпись является первой в мире. Сделана она в 18-м тысячелетии до н.э. и означает следующее:
ama+BARAdara +su-kud-Sin, что в переводе с шумерского означает: «Ама-терасу осуждена Сином». Кифишин расшифровал петроглифы десяти пунктов Восточной Сибири(Прибайкалья). На Рис. 4 приведен петроглиф со скалы у Хана-Шуулун, который Кифишин расшифровал как ama-inanna-BARA2 dara-si и перевел: Ама-терасу (судима) Инанной. Полностью все десять петроглифов Восточной Сибири, расшифрованные Кифишиным, приводятся в статье (18).Авторы считают эти места древними святилищами богини Ама-терасу, принадлежащей к пантеону древних обитателей Север-ной Азии (Сибири), являвшихся предками как шумеров и хеттов-(древних европеоидов), так и японцев (монголоидов). Оказалось, что существуют параллели в пантеонах богов древних шумеров и хеттов, и в японской религии синто. Синтоистская религия японцев –совокупность представлений о существовании богов в виде животных, растений, камней и т.д. Одни лингвисты относят японский язык к алтайской группе языков, другие — к австралийской языковой семье. Но согласно исследованиям Кифи-шина, праяпонцы, как и прашумеры, жили в регионе Алтая и Забайкалья(Селенги), где существовал культ Ама-барагеси. Ама-барагеси упоминается в надписях 2-й Бородинской скалы под Челябинском на Урале (18 000 лет до н.э.), в протошумерском архиве Каменной Могилы в Приазовье (12-3 тыс. лет до н.э.), в архиве месопотамского Ура (3-е тысячелетие до н.э., и в надписях Урнанши из Лагаша (2450 г до н.э.). Итак, мы можем видеть несколько пунктов многотыся-челетней миграции древних протошумеров из региона Восточной Сибири через Приазовье в Переднюю Азию.
В книге индийского ученого Тилака «Арктическая родина в Ведах» приводятся данные из эпических произведений европеоидов Азии о том, что миграция предков индийцев из Циркумполярной зоны шла западнее Урала с севера на юг, а миграция предков иранцев — в том же направлении, но восточнее Урала. Таким образом, как биологические данные — генетические, антропологические, палеозоологические, так и данные археолого-культурологические говорят о том, что европеоиды Европы и Азии, так же, как северные монголоиды и американские индейцы имели центром своей дифференциации центр Азии (Южную Сибирь и соседние регионы), и попали в места своего нынешнего пребывания в результате многотысячелетней миграции.
Выводы.
1. Генетические расстояния человеческих популяций Европы, Азии, Америки, Африки и Океании, построенные по ним дендрограммы, а также распределение гаплогрупп митохондриальной ДНК в человеческих популяциях говорит о том, что европеоиды, так же, как и северные монголоиды, жили в палеолите в Азии, в Сибири, и пришли в места своей современного обитания в результате многотысячелетней миграции.
2. Ряд данных антропологии и палеозоологии поддерживает эту гипотезу палеоазиатского происхождения северных монголоидов, европеоидов и предков американских индейцев.
3. Недавняя расшифровка учеными-лингвистами зачатков праписьменности в Прибайкалье также говорит об обитании европеоидов в палеолите в Сибири, и о древней близости их к монголоидам.
ЛИТЕРАТУРА
1.А.Ф.Назарова, М.Г.Кузнецова. Генетическая структура популяций алтайцев. Доклады РАН, 1993, 333, 3, 405-409.
2.Nazarova A.F.Genetic Data concerning the Problem of Differentiation of Nothern Mongoloids, American Indians and Caucasoids in the Northern Territory of Eurasia. Anthropologisher Anzeiger, 2005, 63,№4, 353-364.
3.M. Nei. The theory of genetic distance and evolution of human races. Jap. J. Human Genet., 1978, v. 23, 341-369.
4.Деренко М.В.,Денисова Г.А., Малярчук Б.А., Дамбуева И.К., Лузина Ф.А., Лотош Е.А., Доржу Ч.М., Карамчакова О.Н., Соловенчук Л.Л., Захаров И.А. Структура генных пулов этнических популяций Алтае-Саянского нагорья на основании данных о поли-морфизме митохондриальной ДНК. Генетика,2001, № 10, 1402-1410.
5.Lahermo P., Sajantila A., Sistonen P., Lukka M.,Aula P.,Peltonen L.,Savontaus M.-L. Amer.J.Hum.Genet.,1996, 58, P.1309-1322.
6.Сукерник Р.И., Шур Т.Г., Стариковская Е.Б. Уоллес Д.С. Изменчивость митохондриальных ДНК у коренных жителей Сибири в связи с реконструкцией эволюционной истории американских индейцев. Генетика, 1996,32, № 3, с.432-439.
7.Brown M.D., Hosseini S.H., Torroni A. et al. mt DNA haplogroup X: an ancient link between Europe/ Western Asia and North America? Amer.J.Hum.Genet.,1998, 63, P.1852-1861.
8.Верещагин Н.К. Вопросы териологии. Общая и региональная териогеография. М., Наука, 1988, с. 5-32.
9.Кобельт В. Географическое распространение животных в холодном и умеренном поясах Северного полушария. СПБ, 1903.
10. А.Ф.Назарова, С.М.Алхутов. Генетические расстояния 55 человеческих популяций Европы, Азии, Америки, Африки и Океании по 28 аллелям 12 локусов белков, ферментов и групп крови. В кн.: Эволюция человеческих популяций, М., Полимедиа, 2000, с.14-20.
11.А.Ф.Назарова. Популяционная генетика русских: генеалогический анализ, частоты генов и генетические расстояния. Доклады РАН, 1994, 339,№ 4, с.563-568.
12.Рычков Ю.Г., Спицын В.А., Шнейдер Ю.В., Назарова А.Ф., Боева С.Б, Новорадовский А.Г., Тихомирова Е.В. Генетика таежных охотников-оленеводов Средней Сибири. Биохимические маркеры генов HP, TF, GC, Alb, GLO I, PGM I, AcP и Es D.Генетика, 1984, т.20, № 10, с.1701-1707.
13.Алексеева Т.И. Этногенез славян по данным антропологии. Изд-во МГУ, 329 с.
14. Дьяконов И.М., Старостин С.А.. (Diakonov I.M., Starostin S.A. Hurro-Urartian as a Eastern Caucasian Language. 1987, Munchen.)
15. Назарова А.Ф. Генетика популяции талышей Азербайджана. Доклады АН СССР, 1991, 317, № 6, с.1484-1486.
16.Алексеев В.П. Палеоантропология земного шара и формирование человеческих рас. М., Наука, 1978, 284 с.
17. Томашевич Т.В. Закономерности распределение частот надглазничных каналов черепа человека. Вопр. Антропол,1988, вып. 80, с.119-128.
18. Кифишин А.Г., Кикешев Н.И. Десять синтоистских святилищ в Прибайкалье. Вестник Международного славянского института, 2005, № 9, 86-94.
Таблица 1. Матрица генетических расстояний между 11 популяциями Европы, Азии и Америками Азии и Америки
|
Популяции |
Генетические расстояния (d) по 28 аллелям 12 локусов (HP, CG, TF, GLOI, Es D, PGMI, ABO, Rh, ACP, AK, ADA, 6-PGD | |||||||||
|
Русские |
– |
|||||||||
|
Немцы |
0,16482 0,03889 |
– |
||||||||
|
Финны |
0,14687 0,03689 |
0,11616 0,03303 |
– |
|||||||
|
Алтайцы |
0,35705 0,05416 |
0,31435 0,05147 |
0,30960 0,05117 |
– |
||||||
|
Эвенки |
0,31725 0,5166 |
0,21287 0,04361 |
0,23986 0,04594 |
0,22245 0,04446 |
– |
|||||
|
Якуты |
0,31907 0,05178 |
0,23037 0,04515 |
0,23837 0,04582 |
0,16566 0,03898 |
0,21568 0,04387 |
– |
||||
|
Монголы |
0,38358 0,05568 |
0,38417 0,05571 |
0,30918 0,05113 |
0,12517 0,03425 |
0,21825 0,04409 |
0,15273 0,03756 |
– |
|||
|
Ненцы |
0,19085 0,04155 |
0,21972 0,04422 |
0,19233 0,04169 |
0,18183 0,04066 |
0,19063 0,04153 |
0,23633 0,04565 |
0,26179 0,04770 |
– |
||
|
Нгасаны |
0,48496 0,06061 |
0,47566 0,06021 |
0,44705 0,05092 |
0,27611 0,04878 |
0,14047 0,03614 |
0,14149 0,03626 |
0,23349 0,04933 |
0,21871 0,04414 |
– |
|
|
Лопари |
0,38710 0,05587 |
0,43152 0,05817 |
0,39181 0,05647 |
0,31477 0,05150 |
0,40745 0,05696 |
0,39380 0,05624 |
0,41203 0,05720 |
0,37814 0,05538 |
0,51185 0,06171 |
– |
|
Америнды |
0,62544 0,06556 |
0,54092 0,06288 |
0,48813 0,06072 |
0,48891 0,06082 |
0,32707 0,05446 |
0,36214 0,05446 |
0,48131 0,06046 |
0,50184 0,06131 |
0,34998 0,05373 |
0,67137 0,06678 |
|
Популяции |
Русские |
Немцы |
Финны |
Алтайцы |
Эвенки |
Якуты |
Монголы |
Ненцы |
Нгасаны |
Америнды |
Таблица 2.
Распределение частот надглазничных каналов черепа человека
|
Популяции |
Частота надглазничных каналов,% |
Популяции |
Частота надглазничных каналов,% |
|
Поляки |
21,3 (47) |
Русские |
35,9 (204) |
|
Армяне |
30,6 (242) |
Манси |
32,1 (112) |
|
Кеты |
33,3 (38) |
Юкагиры |
38,1 (42) |
|
Осетины |
39,4 (314) |
Башкиры северные |
40,0 (122) |
|
Саамы |
47,4 (221) |
Индейцы |
50,2 (124) |
|
Чукчи береговые |
57,1 (70) |
Эвенки |
57,5 (40) |
|
Эскимосы |
57,4 (302) |
Якуты |
63,0 (144) |
|
Англичане |
17,0 (186) |
Монголы |
48,3 (60) |
|
Словене Новгородской губ.,XII-XIV |
32,5 (152) |
Поляне киевские, IX-XIII в.в. |
25,0 (92) |
|
Этруски, VIII-III в.в. до н.э. |
26,9 (70) |
Голландцы |
21,2 (170) |
|
Древние египтяне |
28,3 |
Негры Руанды |
33,6 (61) |
|
Индийцы Уттар Прадеш |
25,2 (238) |
Негры США |
22,3 (202) |
|
Японцы |
43,2 (1008) |
Корейцы |
46,1 (660) |
|
Австралийцы |
19,0 (122) |
Китайцы |
46,5 (202) |
Рис. 1. Микроэволюционная дендрограмма 11 человеческих популяций Европы, Азии и Америки.
Рис.2. Схема палеолитических миграций человеческих популяций, маркированных гаплогруппами митохондриальной ДНК согласно работам разных авторов(3,4, 5,6). В кружках латинскими буквами даны обозначения гаплогрупп мт ДНК.
Рис.3. Микроэволюционная дендрограмма 55 человеческих популяций Европы, Азии, Америки, Африки и Океании. (Названия этнических популяций приведены на самом рисунке).
Рис.4. Петроглиф у Хана-Шулуун из Восточной Сибири (Забайкалья), расшифрованный как палеолитическая письменность древних человеческих популяций.
- Рис.1

-
Рис. 2.
1-саамы,,2-ненцы, 3- нганасаны,4-эвенки,5-якуты,6-монголы,7-алтайцы, 8-русские,9-финны, 10-немцы,11-америнды
- рис.3
- Рис. 4.
E-mail: afnazar@yandex.ru
Назарова А.Ф., Биологические, археологические и культурологические доказательства палеоазиатского происхождения северных монголоидов, европеоидов и американских индейцев // «Академия Тринитаризма», М.,
|
|